

Abstract
Det har blitt stadig tydeligere at den romlige fordeling og bevegelse av membran komponenter som lipider og proteiner er viktige faktorer i reguleringen av mange cellulære funksjoner. Imidlertid, på grunn av den raske dynamikk og de små strukturer som er involvert, blir en meget høy spatio-temporal oppløsning som kreves for å fange den virkelige opptreden av molekyler. Her presenterer vi den eksperimentelle protokollen for å studere dynamikken i fluorescently-merket plasma-membran proteiner og lipider i levende celler med høy spatiotemporal oppløsning. Spesielt, tillater denne metode trenger ikke å spore hvert molekyl, men den beregner populasjon adferd ved bruk av alle molekyler i et gitt område av membranen. Utgangspunktet er en rask avbildning av et gitt område på membranen. Etterpå blir en fullstendig spatio-temporal autokorrelasjonsfunksjonen beregnet korrelering ervervede bilder ved å øke tidsforsinkelser, for eksempel hver 2, 3, N repetisjoner. Det er mulig å vise at breddenfor toppen av de romlige autokorrelasjonen funksjons øker ved økende tidsforsinkelsen som en funksjon av partikkelbevegelser på grunn av diffusjon. Derfor passer av serien av autokorrelasjonsfunksjoner gjør det mulig å trekke ut den faktiske protein betyr kvadratisk forskyvning fra imaging (iMSD), her presentert i form av tilsynelatende diffusiviteten vs gjennomsnittlig forskyvning. Dette gir en kvantitativ visning av gjennomsnittlig dynamikken i enkle molekyler med nanometer nøyaktighet. Ved å bruke en GFP-merket variant av transferrin-reseptor (TfR), og en ATTO488 merket 1-palmitoyl-2-hydroksy-sn -glycero-3-phosphoethanolamine (PPE) er det mulig å observere den tid og rom regulering av protein og lipid på diffusjon mikrometer-størrelse membran regioner i mikro-til-milli-sekunders tidsområde.
Introduction
Starter fra den opprinnelige "flytende mosaikk" modell av Singer og Nicolson, har bildet av mobilnettet plasma membran vært kontinuerlig oppdatert i løpet av de siste tiårene for å inkludere den nye rollen cytoskjelett og lipid-domener 1,2.
De første observasjonene ble innhentet av fluorescerende gjenoppretting etter fotobleking (FRAP) avsløre at en betydelig andel av membran proteiner er immobile 3-5. Disse banebrytende studier, men veldig lærerikt, led av den relativt dårlig oppløsning i verdensrommet (mikron) og (i sekunder) av FRAP oppsett. Også, å være et ensemble gjennomsnittsmåling, mangler FRAP i å gi informasjon om enkelt molekyl atferd.
I denne sammenheng er det mulig å spesifikt merke et enkelt molekyl med svært lyse koder (som tillater studiet av diffusjonsprosess ett molekyl av gangen) har vært meget vellykket. Spesielt, ved å skyvetidsoppløsning av Single Particle Tracking (SPT) tilnærming til mikrosekunder tidsskala, Kusumi, et al. fått tilgang til ukjente funksjoner i lipid og protein dynamikk som i stor grad bidratt til anerkjennelse av den rollen aktin-basert membran skjelett i membran fysiologi 6 7. Disse funnene ble generert i så-kalt «plukket og gjerdet 'modellen, der lipid og protein diffusjon reguleres av aktin-baserte skjelett. Men, for å ha tilgang til den enorme mengden av informasjon gitt av SPT mange eksperimentelle saker må tas opp. Spesielt er merkningsmetode vanligvis består av mange trinn som produksjon, rensing og innføring av de merkede arter inn i systemet. Videre store etiketter, som quantum prikker eller metallnanopartikler, er ofte nødvendig for å nå den undermillisekund tidsskala, og tverrbindingen av målmolekylene av etiketten ikke kunne unngås i mange tilfeller. Til slutt, mange banerhar å bli tatt opp for å passe statistiske kriterier og samtidig en lav tetthet av etiketten er nødvendig for å tillate sporing.
Sammenlignet med SPT, fluorescens korrelasjonsspektroskopi (FCS), å overvinne mange av disse ulempene, utgjør en meget lovende tilnærming for å studere molekyldynamikk. Faktisk fungerer FCS vel også med dim og tette etiketter, slik at å studere dynamikken i fluorescerende protein-merket molekyler i forbigående transfekterte celler. Dessuten gjør det nå høye statistikken i en begrenset tidsperiode. Endelig er, til tross for "høy" densitet på etiketter FCS gir enkle molekyler informasjon. Takk til alle disse egenskapene, FCS representerer en veldig grei tilnærming og har blitt mye brukt til å studere lipid og protein dynamikk både i modellmembraner og i levende celler 8-10. Mange forskjellige metoder er blitt foreslått for å øke evnen til FCS for å avdekke detaljer av molekylær diffusjon. For eksempel, det var shegne som ved å utføre FCS på forskjellig størrelse observasjon områder kan man definere en "FCS diffusjon lov" opplysende skjulte funksjoner i molekylær bevegelse 11,12. Dessuten blir variert i størrelse, ble det sentrale område også duplisert 13, beveges i rommet langs linjene 14-20 eller konjugert med raske kameraer 21,22. Ved hjelp av disse "spatio-temporal 'korrelasjon tilnærminger, ble relevante biologiske parametere av flere membran komponenter kvantitativt beskrevet på både modellmembraner og faktiske biologiske seg, noe som gir innsikt i membran romlige organiseringen.
Men i alle de FRAP FCS og anvendelser som er beskrevet så langt størrelsen på det sentrale området utgjør en begrensning i romlig oppløsning som ikke kan overvinnes. Flere super-Resolution Imaging metoder har nylig blitt utviklet for å omgå denne begrensningen. Noen er basert på lokalisering presisjon, for eksempel stokastisk optisk gjenoppbygging mikroskopi (STORM) <sup> 23,24, fotoaktivering lokalisering mikroskopi (PALM) 25, fluorescens PALM (FPALM) 26, og enkeltpartikkelsporings PALM (sptPALM) 27: den relativt store mengden av fotoner som kreves ved hvert stillbilde, men begrenser tiden oppløsningen disse metodene til minst flere millisekunder, og dermed hindre deres anvendelse i vivo.
I kontrast, har et lovende alternativ for oppløsning super blitt åpnet av romlig moduler fluorescensemisjonen med stimulert emisjon uttømming metoder (STED eller reversible metningsbar optiske fluorescens overganger (RESOLFT)) 28,29. Disse tilnærmingene kombinere utformingen av observasjons volum godt under diffraksjon grensen med muligheten til å bruke rask skanning mikroskoper og deteksjonssystemer. I kombinasjon med fluorescens svingninger analyse, STED mikros lov til å direkte sondere nanoskala tid og rom dynamikken i lipider og proteins i levende cellemembraner 30,31.
De samme fysiske mengder STED-baserte mikroskopi kan oppnås ved en modifisert spatio-temporal bildekorrelasjonsspektroskopi (pinner 32,33) metode som er egnet for undersøkelsen av dynamikken i fluorescensmerket-merkede membranproteiner og / eller lipider i levende celler og ved en kommersielt mikroskop. Den eksperimentelle protokollen som presenteres her er komponert av noen få trinn. Den første krever en rask avbildning av regionen av interesse på membranen. Deretter blir den resulterende stabel av bilder som brukes for å beregne den gjennomsnittlige romlige-tidsmessige korrelasjonsfunksjoner. Ved montering av serien av korrelasjonsfunksjoner, kan den molekylære 'diffusjon lov »fås direkte fra avbildning i form av en tilsynelatende diffusivitet (D app) - vs -average forskyvning plot. Denne tomten kritisk avhengig av miljøet utforsket av molekylene og lar erkjenner direkte selve diffusjon moduserav lipid / protein av interesse.
I flere detaljer, som tidligere er vist 34, den spatio-temporal autokorrelasjonsfunksjonen til ervervet bildeserien kritisk avhengig av dynamikken i molekylene beveger seg i samlet bildeserien (bemerk at den samme resonnement kan anvendes i en linje anskaffelse der bare én dimensjon i verdensrommet er vurdert). Spesielt definerer vi korrelasjonsfunksjon som:
 (1)
(1)
hvor  representerer den målte fluorescensintensiteten i posisjon x, y, og ved tidspunktet t,
representerer den målte fluorescensintensiteten i posisjon x, y, og ved tidspunktet t, ![]() og
og ![]() representerer avstanden i x ogy-retningene henholdsvis
representerer avstanden i x ogy-retningene henholdsvis ![]() representerer tidsforsinkelsen, og
representerer tidsforsinkelsen, og ![]() representerer gjennomsnittet. Denne funksjon kan uttrykkes som:
representerer gjennomsnittet. Denne funksjon kan uttrykkes som:
 (2)
(2)
hvor n representerer det gjennomsnittlige antall molekyler i observasjonsområdet, ![]() representerer konvolusjon drift i rommet, og
representerer konvolusjon drift i rommet, og  representerer autokorrelasjon av instrumental midjen. Dette siste kan tolkes som et mål på hvordan fotoner av en enkelt emitter er spredt ut i rommet på grunn av den optiske / opptaksoppsettet (den såkalte punktspredefunksjon, PSF, genetrally godt rundet av en Gaussisk funksjon). Endelig
representerer autokorrelasjon av instrumental midjen. Dette siste kan tolkes som et mål på hvordan fotoner av en enkelt emitter er spredt ut i rommet på grunn av den optiske / opptaksoppsettet (den såkalte punktspredefunksjon, PSF, genetrally godt rundet av en Gaussisk funksjon). Endelig  representerer sannsynligheten for å finne en partikkel i en avstand
representerer sannsynligheten for å finne en partikkel i en avstand ![]() og
og ![]() etter en tidsforsinkelse
etter en tidsforsinkelse ![]() . Hvis vi vurdere et diffusiv dynamikk, der partiklene beveger seg tilfeldig i alle retninger og netto fluks ikke er til stede, er denne funksjonen også godt rundet av en Gaussisk funksjon der variansen kan identifiseres som Mean Square Vekt (MSD) av bevegelige partikkel . Således midje av korrelasjonsfunksjon (også omtalt som
. Hvis vi vurdere et diffusiv dynamikk, der partiklene beveger seg tilfeldig i alle retninger og netto fluks ikke er til stede, er denne funksjonen også godt rundet av en Gaussisk funksjon der variansen kan identifiseres som Mean Square Vekt (MSD) av bevegelige partikkel . Således midje av korrelasjonsfunksjon (også omtalt som ![]() ), Kan defineres som summen av de partikkel MSP og den instrument midje og kan måles ved en gaussisk formlingen av korrelasjonsfunksjonen for hvert tidsforsinkelse. Den måles i MSD kan brukes til å beregne en tilsynelatende diffusivitet for de bevegelige molekyler
), Kan defineres som summen av de partikkel MSP og den instrument midje og kan måles ved en gaussisk formlingen av korrelasjonsfunksjonen for hvert tidsforsinkelse. Den måles i MSD kan brukes til å beregne en tilsynelatende diffusivitet for de bevegelige molekyler ![]() og en gjennomsnittlig forskyvning
og en gjennomsnittlig forskyvning ![]() som:
som:
 (3)
(3)
 (4)
(4)
Noen betraktninger om den eksperimentelle oppsettet brukt kan veilede leseren gjennom de følgende avsnittene. For selektivt å eksitere fluoroforer i basalmembranen av levende celler vil vi bruke en total intern refleksjon (TIR) belysning, ved hjelp av en kommersiell TIR fluorescens (TIRF) mikroskop (detaljer kan finnes i materialet avsnitt). Videre, for å samle the fluorescens vil vi bruke en høy forstørrelse objektiv (100X NA 1,47, er høy numerisk apertur som kreves for TIRF belysning) og en EMCCD kamera (fysiske størrelsen på pixel på brikken 16 mikrometer). For å nå en pikselstørrelse på 100 nm vi bruke en ekstra forstørrelse linsen på 1.6x. Som omtalt nedenfor, vil en tidsoppløsning under 1 ms være nødvendig å beskrive riktig dynamikken i membranlipider raske under 100 nm. For å nå dette tidsoppløsning må vi velge et område av interesse (ROI) mindre enn hele chip av kameraet (512 x 512). På denne måten, vil kameraet lese et redusert antall linjer øker tidsoppløsning. Men i denne avlesning regime rammen tid ville være begrenset av tiden det tar å skifte summen fra eksponering for avlesning chip på kameraet og er vanligvis i størrelsesorden millisekunder for 512 x 512 pixel EMCCD. Å slå denne grensen, kan en ny teknologi skiftende ROI-linjer bare i stedet for hele rammen, wed en praktisk effektiv reduksjon av chip størrelse eksponert (kalt Cropped Sensor Mode i vår EMCCD). For denne konfigurasjon for å være effektiv, må chip utsiden av ROI dekkes med et par slisser som er montert i den optiske banen. Takket være denne sette opp en tidsoppløsning ned til 10 -4 sekunder kan oppnås. Vær imidlertid oppmerksom på at denne tilnærmingen kan være kombinert med mange ulike forsøksoppsett, som forklart i 'diskusjon' del.
Demonstrasjon av fremgangsmåten vil bli gitt i levende celler, ved å bruke både en ATTO488 merket 1-palmitoyl-2-hydroksy-sn -glycero-3-phosphoethanolamine (ATTO488-PPE) og et GFP-merket variant av transferrinreseptoren (GFP- TfR). I tilfelle av ATTO488-PPE denne tilnærmingen kan med hell gjenvinne en nesten konstant D app som en funksjon av gjennomsnittlig forskyvning indikerer en hovedsakelig fri diffusjon, som tidligere rapportert 30,35. Derimot, viser TfR-GFP en avtagende D
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. systemkalibrering
- Punktspredefunksjon (PSF) kalibrering
- Fortynn 10 mL av 30 nm fluorescerende bead-løsning (omtrent 5 mM) i 90 mL av destillert vann og deretter sonicate løsningen i 20 min. Skjær en firkant (1 cm x 1 cm) stykke agarosegel (3%) og innskudd 10 ul av oppløsningen på toppen av gelen. Velte stykke gelen i bunnen av glass en 2 cm petriskål og klem fallet på glasset.
- Slå på kjøpet oppsett, sette prøven i holderen, sett kameraet eksponering og EMgain (100 msek og 1000 er gode parametre, men optimalisere henhold til systemet) og vente på at kameraet skal kjøle seg ned.
- Sett kameraet eksponeringen til 100 msek, kamera EMgain til 1000, oppkjøp modus til Frame Transfer, 100 repetisjon og automatisk lagre innstillingen.
- Ved hjelp av okularet og overført lys fokus på grensen av gel og deretter flytte målet til midten av gel, justere fokus og start laser justering prosedyre (i LAS AF, velg 'TIRF setup "og følg den automatiske justeringsprosedyren).
- Finn et synsfelt med isolerte enkelt flekker, nøyaktig fokusere på den lysere flekk (som vanligvis representerer perler aggregat) som en referanse, erverve 100 bilder og gjenta trinn 5-6 ganger for å skaffe flere enkelt flekker.
- Importere kjøpt serien til en databehandler og gjennomsnittlig stabelen i tid (figur 1A) og velge en enkelt isolert perle. Vær nøye med å velge de minste å unngå partikkel aggregater.
- Monter den valgte intensitetsfordeling (et eksempel på enkelt perler profilen er presentert i figur 1B) med en Gauss-funksjon ved hjelp av kommandoen "gaussfit" (i ICS-Matlab verktøy i materialet i Matlab). Verifisere godhet av passform ved å inspisere de oppnådde residualene (et eksempel av montert Gaussian profilen med de tilsvarende residualene er presentert in Figur 1B).
- Kamera kalibrering
- Slå på kameraet og vente på at kameraet skal kjøle seg ned. Still kamera oppkjøpet innstilling, (dvs. for det brukte kameraet vi satt eksponeringen til 0,5 msek, kamera EMgain til 1000, oppkjøp modus til Cropped Mode, ROI størrelsen til 32 x 128, 10 000 repetisjoner) og starte kjøp av kameraet bakgrunn signal.
- Import kjøpt ramme serien til en databehandler. Beregn og inspisere den gjennomsnittlige intensitet i hver piksel for å verifisere at kameraet bakgrunnen er tilnærmet flat i det valgte område av brikken. På nav-modus, fjerner den første og de siste få horisontale linjer (3 til 10, avhengig av størrelsen på ROI) for hver ramme fordi kameraet bakgrunnen er vanligvis forspent i grenselinjer.
- Lag et histogram av verdiene (også definert Digital Level, DL) i kjøpte bilder stakken (ved hjelp av kommandoen "hist" i Matlab) og plotte logaritmenav resulterende frekvens (ved hjelp semilogy kommando i Matlab). Et eksempel på DL fordeling for kameraet bakgrunn er presentert i figur 2.
MERK: Hvis kameraet fungerer godt, vil plottet viser en tilnærmet Gaussisk peak (en parabolsk profil i log-skala) som representerer fordelingen av verdier knyttet til null foton etterfulgt av en eksponentiell (en linje med negativ helning i log-skala ) som representerer fordelingen av verdier knyttet til en foton (figur 2). Spesielt midten og variansen av den gaussiske funksjonen representerer kamera offset og feiling, henholdsvis, mens nedbrytningskonstanten for den eksponensielle delen utgjør en estimering av DL tildelt av kameraet til hver enkelt foton. I Matlab bruker avsnittet "CalibrateCamera" av Script i støttemateriell. - Gjenta operasjonen for alle de valgte kameraet EMGain og Gain.
2. MerketCell Forberedelse
- For å fremstille liposomer som er nødvendige for lipid inkorporering 36, oppløses separat 1 mg av DOPE (1,2-dioleoyl- sn -glycero-3-phosphoethanolamine), 1 mg DOTAP (1,2-dioleoyl-3-trimetylammonium-propan), og 1 mg av PPE-ATTO488 i en ml kloroform. Bland 0,5 ml av DOPE-løsning, 0,5 ml av DOTAP-løsning, og 25 pl av PPE-ATTO488 løsning og tørr under vakuum i 24 timer. Tilsett 0,5 ml av HEPES-buffer 20 mM, vortex i 15 min og sonicate i 15 min ved 40 ° C.
- For å forberede cellen, vask 3 ganger med PBS en p100 rett av konfluent CHO-K1 (Chinese Hamster eggstokk), tilsett 1 ml trypsin og butikk i inkubator for 5 min. Suspender delte celler legge 9 ml DMEM / F12 medium supplert med 10% FBS og frø 150 ul celle-løsning i en petriskål inneholdende 800 pl av det samme mediet.
- Oppbevares i inkubator i 24 timer ved 37 ° C og 5% CO 2. For lipid innlemmelse, bytte celle medium med 500mikroliter av serumfritt medium; etter 30 min, tilsett 2 mL av liposomer løsning; etter 15 min vask med PSB og legge til nye DMEM / F12 medium for bildebehandling.
- For Transfeksjon, transfektere celler ifølge Lipofectamine protokollen (Produsent instruksjoner) bruker TfR-GFP plasmid og lagre 24 timer i kuvøse før bildebehandling.
3. Data Acquisition
- Setup forberedelse
- For å termostat mikroskopet, 24 timer før forsøket slå på inkubator.
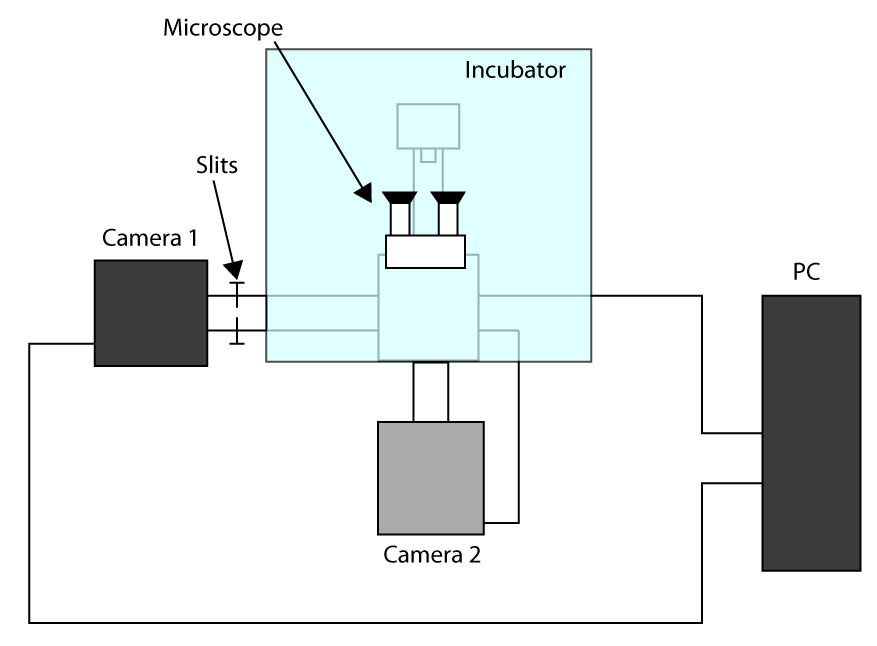
- For å søke raskest mulig oppkjøp tid, arbeid i Cropped Sensor Mode (se innledningen) og bruke et første kamera for imaging (kamera 1) og andre kameraet for å velge cellen (kamera 2). En ordning av konfigurasjon er presentert i Tilleggs Figur S1. Så, for å justere to kameraer slå på mikroskopet og vente på kameraene for å kjøle seg ned.
- Sett på begge kameraene parametrene for fallende lys imaging (dvs.
- Sett prøvene i holderen og fokusere med okular, sende lys til kamera 1 og skyv spaltene slik at lyset bare på ROI brukes for celle imaging (her en 32 x 32 piksler ROI).
- Flytt en celle i det valgte område og sender lyset til kameraets 2, og deretter trekke et ROI i programvaren som styrer kamera 2 for å få en referanse.
- Imaging (figur 3A)
- Først av alt, må du justere TIRF laser i henhold til prosedyren for ditt oppsett. Velg 'TIRF setup "i vår oppsettet og starte automatisk justering prosedyren. Når laseren er på linje satt 70 nm av inntrengningsdybden (ca. 70 °).
- Sett eksponeringstid til 70 msek og EMGain til 100 på både kamera 1 og kamera 2; deretter velge en celle ved hjelp av kamera 1, deretter sende lyset på kamera 2 og nøyaktig fokus cellemembranen. Sette minimum eksponering på camera 2, 1000 EMGain, Cropped Sensor Mode, 10 5 repetisjoner og sett autolagring som passer filer (Fleksibel bilde Transport System, et format som enkelt kan styres).
- Starte kjøp å registrere bildeserie. Slipp Gain og det beskårne Mode for å tillate temperaturstabilisering før du henter en ny celle, og deretter gjenta de to siste trinnene for å tilegne seg 8-10 celler.
4. Beregning av Mean Square Displacement fra Imaging (i MSD)
MERK: Følgende protokoll kan direkte brukes til rådata. Samtidig er hele protokollen gjelder for datainnsamlings- simulert både i Matlab og i SimFCS. Linken til de tilsvarende tutorials kan bli funnet i "Materials 'delen.
- Beregning av Matlab
- Importere kjøpte serien i Matlab ved hjelp ImportImageSeries script. Beregn den gjennomsnittlige intensitet av hvert bilde i tid ved hjelp av command mener på de første to dimensjoner og bruke tomten for å se den resulterende vektor.
- Dersom mer enn 10% av fotobleking er til stede, skal den kastes serien eller fjerne første del av dem. Hvis den er lavere, kan du prøve å rette effekten på korrelasjonsfunksjonen ved å trekke til hvert bilde sin middels intensitet, som vist før 37.
- Beregn den gjennomsnittlige intensitet av hver piksel ved å benytte middelverdien i tredje dimensjon og se resulterende bildet.
MERK: Spesiell oppmerksomhet er nødvendig for å unngå kunstig sammenhenger. Faktisk, som tidligere vist for lignende teknikker 38, cellekantlinjer så vel som ute av fokus vesikler kunne introdusere en sterk korrelasjon. Hvis inspeksjon av gjennomsnittlig bildet avslører cellekantlinjer eller ute av fokus vesikler, prøv å utelukke regionen involvert ellers forkaste oppkjøpet. For å korrigere for dette immobile strukturer trekke den gjennomsnittlige time intensiteten fra hver piksel 39. - Beregn than spatiotemporal korrelasjon (G (ξ, χ, τ)) ved å benytte funksjonen CalculateSTICScorrfunc. Fjern G (ξ, χ, 0), fordi korrelasjonen skyldes skuddet støy i dårlig lys regime dominerer G (0,0,0); korrelasjonen grunn av detektoren dominerer G (± 1,0,0), og partikkelbevegelsen i løpet av eksponeringstiden kunne deformeres G (ξ, χ, τ) for τ = 0 ved å øke den målte midjen (denne effekten forsvinner for τ > 0) 34.
- Gjennomsnittlig G (ξ, χ, τ> 0) under anvendelse av en logaritmisk tids-bin for å redusere støy ved å bruke "LogBinStack" funksjon i Støttemateriale og deretter passe den resulterende G (ξ, χ, τ) ved hjelp av funksjonen "gaussfit" i ICS-Matlab verktøy i materialet for å gjenopprette i MSD (den andre kolonnen i den resulterende matrisen).
- Plott erholdt midje σ (τ) 2 (i MSD) som en funksjon av tiden. Hvis dataene er for bråkete, prøv å øke antall oppkjøpet soIRED rammer, øke lasereffekten, gjennomsnittlig mer G (ξ, χ, τ) sammen.
- Beregning av SimFCS
- Åpne de ervervede filer med ImageJ hjelp BioFormat importør plugin og lagre ervervet serie som Tiff sekvens.
- Åpne SimFCS og velg RICS verktøyet og velg Fil> Importer flere bilder (Supplementary Figur S2).
- Velg Fit, setter de riktige oppkjøps parametere og lukke fit vindu (Supplementary Figur S3).
- Velg Display> middels intensitet> CH1 og bekrefte tilstedeværelse av fotobleking (Supplementary Figur S4).
- Hvis mer enn 10% av fotobleking er til stede forkast serien, eller hvis det er mulig belastning på nytt bilde fjerne den første delen av serien sekvensen.
- Hvis bleking den er lavere enn 10% velger du Verktøy> i MSD> sette parametre, sjekk 'Bruk glidende gjennomsnitt ", satt i ROI panelet til venstre enumbra av rammen for det glidende gjennomsnittet å være oppmerksom på at abonnenten er høyere enn den karakteristiske diffusjon tid (for partikkel beveger seg i en mikrometer 2 sek -1 en tid på 10 sekunder er et godt glidende gjennomsnitt)
- Velg Verktøy> iMSD> Beregn Jeg MSD (Supplementary Figur S5) og passform og eksportere i MSD fra memo pad (Supplementary Figur S6).
5. Beregning av Diffusion Law fra i MSD
- Monter de første par punkter å ekstrapolere skjærings (σ 0 2) (5 poeng er vanligvis nok, men flere poeng kan monteres hvis de viser en lineær oppførsel) og sammenligne denne verdien med den tidligere målt PSF to. Hvis de er sammenlignbare, er dynamikken til isolerte fluoroforer følges. Derimot, hvis σ 0 2 >> PSF 2 prøve å tilegne seg raskere for å sikre atingen skjulte dynamikken er tilstede 34.
- Beregn den tilsynelatende diffusivitet (D app) og gjennomsnittlig forskyvning (R) ved hjelp av ligningene 3 og 4 (se innledningen).
- Plot D app som en funksjon av R for å oppnå en diffusjon lov sammenlignbar med det som er målt med flekk variasjon basert FCS 12 (figur 3D).
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
For å kalibrere den instrument midjen, kan bildet av en enkelt fluorescerende nano-vulsten være tiltak som beskrevet i protokoll trinn 1.1. En typisk fluoriserende bilde av disse perlene er presentert i figur 1. Festingen av intensitetsfordelingen av en 2D gaussisk funksjon gir tilbake gode restprodukter, og gjør det mulig å måle den instrument midjen på 270 nm. Denne verdien er i god overensstemmelse med den forventede diffraksjon grensen anslått av Rayleigh ligningen. Denne kalibrering ikke er nødvendig for måling av partikkel-dynamikk, men det er nødvendig å måle den tilsynelatende partikkelstørrelse.
En typisk frekvensfordeling av kameraet bakgrunn er presentert i figur 2. Toppen ved ca 180 DL er avhengig av kameraets respons på ingen foton, og det representerer bidraget av Analog digital (AD) omformere. Dette bidraget kan tilnærmes som en gaussisk fordeling å anslå offset og varianseninnført av signalopptak. Over 200 DL det digitale nivå fordelingen blir eksponensiell (lineært i logaritmisk skala) og som representerer den gjennomsnittlige kameraet respons på en enkelt foton. Montering av denne del med en eksponensiell fordeling tillater målinger av gjennomsnittlig DL tildelt hver enkelt foton. Jo høyere er forholdet mellom den gjennomsnittlige DL tilordnet hver foton og AD-omformer feil, vil den nedre være støy i den beregnede korrelasjonsfunksjonen. Videre tillater den gjennomsnittlige enkelt foton respons på estimeringen av kameraets dynamiske område.
Et diagram av den fullstendige eksperimentelle prosedyren er oppsummert i figur 3, og et bilde av Atto488-PPE innsetting inn i membranen er representert i figur 4A. Et representativt TIRF bilde av basalmembranen av en CHO-celler merket med Atto488-PPE er presentert i figur 4B. Flere lyspunkter kan være til stede utenfor cellen på grunn av liposomes stablet på glasset. De kan kastes ved å velge en avkastning på et membranparti stort sett ensartet i fluorescens (f.eks. Den cellulære plasmamembranen). Som forventet målt diffusjon lov (figur 4C) for dette lipid er flat, noe som indikerer et stort sett gratis diffusjon som tidligere vist ved STED-FCS målinger 30,35. Det er verdt å nevne at alle de viste forskyvningsverdiene er under diffraksjon grense, som klart viser evnen av denne tilnærmingen til super-løse midlere molekyl forskyvninger godt under grensen diffraksjon og ned til noen få titalls nanometer.
En skjematisering av TfR-GFP dimer innsetting inn i membranen er representert i figur 5A. Mange studier viste at den cytoplasmiske hale av denne reseptoren interagerer med membranen skjelett, som i sin tur virker som et gjerde for reseptoren mobilitet 12,40. En representant TIRF bilde av en CHO celle uttrykker TfR-GFP er presented i figur 5B. Lav fluorescensintensitet celler bør foretrekkes, da membranen er nærmere til den opprinnelige tilstand, og sannsynligheten for gjenstander i forbindelse med over-ekspresjon er minimert. I tillegg bør den sentrale del av cellen unngås, da virkningene av ut-av-fokus fluorescens (fra cytoplasma, for eksempel) kan være tilstede. Som forventet er målt diffusjon lov (figur 5C) i TfR-GFP viser en første flat oppførsel under 100 nm, med en gjennomsnittlig D app på ca 0,7 um 2 sek -1, etterfulgt av påfølgende rask reduksjon i tilsynelatende diffusivitet ned til 0,2 mikrometer 2 sek -1 (verdien vanligvis målt ved diffraksjon begrenset FCS 12). Dette resultatet viser at vår tilnærming lett kan måle den gjennomsnittlige forskyvning av GFP-merkede proteiner med en oppløsning på noen få titalls nanometer. Videre den romlige skalaen der D app begynner å minke sett den karakteristiskeromlig skala fra protein delvis innesperring av membranen skjelettet på rundt 120 nm, i tråd med tidligere estimater seks.

Figur 1. Kalibrering av punktspredefunksjon. (B) Pseudo bilde av en isolert perle og perler aggregater. (B) 3D plott av intensitetsprofilen til en isolert vulsten viser en veldefinert gaussisk profil. (C) Fit av intensitetsfordelingen av en Gauss-funksjon (øvre panel) med de tilsvarende rester (nedre panel). Den gode avtalen mellom utstyrt distribusjon og den målte intensiteten profilen er også et bevis på at instrumental PSF kan tilnærmes av en Gaussisk funksjon. Vennligst klikk her for å VI ew en større versjon av dette tallet.

Figur 2. Kalibrering av kamera respons på enkelt fotoner. Figuren viser Digital Level (DL) distribusjon for kamera bakgrunn i en 32 x 128 ROI, eksponering 0,5 msek i Cropped Sensor Mode. Toppen ved ca 180 DL representerer kameraet respons på ingen fotoner. Spesielt, den representerer bidraget fra den analoge digital (AD) omformere, og kan tilnærmes av en gaussisk funksjon å beregne forskyvningen og variansen innført av signalopptak. Over 200 DL fordelingen av digitale nivåer blir eksponensiell og representerer den gjennomsnittlige kameraet respons på en enkelt foton. Målingen av disse parametere tillater å estimere tettheten av fotoner som er registrert under overtakelsen.s / Ftp_upload / 51994 / 51994fig2highres.jpg "target =" _blank "> Klikk her for å se en større versjon av dette tallet.

Figur 3. skjematisering av metoden. (A) Vidvinkelokular avbildning av EMCCD kamera er brukt for å nå sub-millisekund oppløsning, mens TIRF microcopy utnyttes for å gi nøyaktig optisk seksjonering av plasmamembranen. (B) Den resulterende bunke bilder er autokorrelert for å beregne gjennomsnittlig romlig -temporal korrelasjonsfunksjonen. Denne korrelasjonsfunksjon er godt tilnærmet ved en gaussisk funksjon (se innledningen) og det sprer seg ut i tid i henhold til partikkelforskyvninger. (C) Således, for å kvantifisere den spredning av korrelasjonsfunksjonen på grunn av molekyl fortrengning, passende med en Gauss ian funksjonen utføres. Dette gjør det mulig å måle den molekylære 'diffusjon lov' direkte fra bildebehandling, i form av tilsynelatende diffusiviteten vs gjennomsnittlig forskyvning tomten. (D) Takket være denne tomten, kan molekylær diffusjon moduser bli direkte identifisert med ikke behov for en fortolkningsmodell eller antakelser om romlig organisering av membranen. Faktisk vil fritt diffunderer molekyler viser en konstant relativ diffusivitet som deres mobilitet ikke er avhengig av den romlige målestokk av målingen. Derimot, vil delvis trange molekyler vise en ganske konstant relativ diffusiviteten for forskyvninger mindre enn innesperring størrelse, deretter en avtagende diffusiviteten for romlige skalaer større enn innesperring størrelse. Følgelig kan forekomsten av en reduksjon i den tilsynelatende diffusivitet tolkes som et fingeravtrykk av transient sperring, mens den tilhørende romlige målestokk kan brukes til å estimere den romlige utvidelse av innesperring. .jove.com / filer / Ftp_upload / 51994 / 51994fig3highres.jpg "target =" _blank "> Klikk her for å se en større versjon av dette tallet.

Figur 4. ATTO488-PPE diffusjon lov i levende cellemembraner (A) Skjematisk fremstilling av ATTO488-PPE innsetting i cellemembranen (B) TIRF bilde av CHO basalmembranen merket med ATTO488-PPE:.. En ROI (rød boks) er valgt i en stort sett jevn del av cellen, unngå cellekanten og sterkt fluorescerende flekker. (C) Spredningen lov målt i den valgte ROI viser en flat atferd bekrefter en gratis spredningsmodell for denne komponenten. Vennligst klikk her for å se en større versjon av dette tallet.

Figur 5. TfR-GFP diffusjon lov i levende cellemembraner (A) Skjematisk fremstilling av TfR-GFP innsetting i cellemembranen:. Cytoplasmatiske halen av reseptoren samhandler med membranen skjelett, som fungerer som et gjerde for reseptor mobilitet (B. ) TIRF bilde av CHO uttrykke TfR-GFP: en ROI er valgt foretrakk lave uttrykker cellene å unngå gjenstander på grunn av overekspresjon (C) diffusjon lov TfR (svarte prikker), i motsetning til PPE (grå linje, tatt fra figur 4),. viser den typiske oppførselen delvis begrenset diffusjon der en første flate delen er etterfulgt av en nedgang i D app. Vennligst klikk her for å se en større versjon av denne figur.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Enkelt partikkel tracking (SPT) representerer en av de mest vanlige strategier for å studere molekyldynamikk, og det har den store fordelen av å måle partikkelbaner. Dette i sin tur gjør det mulig for sondering oppførselen selv noen merkede partikler i et komplekst system. Imidlertid, for å oppnå denne fordel SPT må vanligvis en lav tetthet av sonden og meget lyse etiketter. Spesielt, for å oppnå høy tidsoppløsning (usekunder range) uorganisk probe er vanligvis nødvendig (f.eks kvanteprikker eller metallnanopartikler): i dette tilfellet er det nødvendig med en komplisert prosedyre for produksjon, merking og innsetting inn i systemet. Sammenlignet med SPT den nåværende metoden viser noen viktige fordeler. Først av alt, kan denne tilnærming brukes i konjugering med fluorescerende proteiner. Dermed sammenlignet med SPT, er en høyere tidsoppløsning oppnådd (på samme etikett) takket være lavere mengde fotoner kreves 34. Mer i detalj, gjør denne eiendommen skyve den time resolution under 10 - 3 sek også når du bruker encodable fluorescerende proteiner, og dette tidsskala gir eksklusiv tilgang til nanoskala dynamikken i membran bestanddeler. Til slutt er det verdt å merke seg at molekylær diffusjon lover beskrives ved å analysere hele rom-tid korrelasjon funksjon, uten behov for å spore hvert molekyl.
Sammenligningen med STED-baserte FCS er også interessant. I en STED-FCS måling gjennomsnittlig overføringstiden av molekyler for å redusere observasjonsvolum måles ved tidsmessig korrelasjon av fluorescens-signalet. Dette gjør det mulig å skaffe en lokal måling av den molekylære dynamikk også under diffraksjon grense. I presenterte tilnærming spredningen loven er målt som gjennomsnittet av alle partiklene beveger seg i den valgte ROI, observert ved hjelp av standard, diffraksjon-begrenset, observasjon volum. Imidlertid rapporterte resultater viser at denne metoden ikke er begrenset av diffraksjon, men bare ved the tidsmessig oppløsning tilgjengelig. Faktisk, selv om en diffraksjon begrenset anskaffelse blir brukt til å detektere fluktuasjoner (analogt til det som er gjort i andre superoppløsningsteknikker, som for eksempel PALM og STORM), molekyl forskyvninger godt under diffraksjon grensen kan være (direkte) beregnet, som allerede demonstrert ved hjelp av pinner for å måle molekyl renner 32. Videre, i motsetning STED-FCS, denne tilnærmingen enkelt kan brukes til et bredt spekter av kommersielle og eksisterende mikros oppsett, for eksempel rasteravsøksmetoder mikroskoper eller bredt felt kamerabaserte mikroskoper. Det er verdt å nevne at STED-FCS målinger av molekylære diffusjon lover strengt krever en Fluoroforen avhengig kalibrering av størrelsen på instrumental midjen. Motsatt, presentert målingen her ikke krever en systemkalibrering (kun nødvendig for estimering av partikkelstørrelse).
Selve oppløsning i måling av partikkel forskyvninger av de presenterte fremgangsmåteavhenger i høy på hvor nøyaktig vi kan måle korrelasjonsfunksjonen. Følgelig er det ikke i seg selv er begrenset av diffraksjon, analogt med SPT tilfelle hvor oppløsningen er avhengig av hvor nøyaktig partikkel "bilde" blir målt. For å måle en signifikant korrelasjon i mindre enn 1 min for de foreslåtte eksperimenter, få fotoner (vanligvis under 10 fotoner) for hver partikkel i hver ramme er nok. Faktisk, er bidraget fra alle de observerte partikler gjennomsnitt sammen når korrelasjonsfunksjon blir beregnet, selv om partiklene ikke er isolert. Denne eiendommen er iboende svingninger korrelasjonsmetoder og lar Bruke dim og tette etikettene, for eksempel fluorescerende proteiner transfektert i levende celler.
Med tanke på dette synes det klart at den minste målbare forskyvning avhenger av diffusivitet til partikkelen, og på tidsoppløsning på bildeoppsettet. Som et eksempel, kan vurdere diffusjon av molekyler på cellemembran,der høyeste målte diffusiviteten for proteiner eller lipider er rundt fem mikrometer 2 sek -1. Under slike forhold trenger vi en tidsoppløsning på ca 10 -4 sek å fange en gjennomsnittlig forskyvning på 50 nm. Denne tidsoppløsning kan oppnås ved rask scanning mikroskop langs enkeltlinjer eller ved hurtig EMCCD kamera, hvor tidsoppløsning faller sammen med eksponeringstiden, som vist her.
En ytterligere vesentlig forutsetning for denne metoden for å beskrive nøyaktig molekylær dynamikk er en korrekt romlig sampling. Faktisk, for å passe korrelasjonsfunksjonen trenger vi en spatial sampling (pikselstørrelse) lavere enn midjen på instrumental PSF. I de fleste kommersielle mikroskoper (konfokal eller bredfelt), spenner over PSF midjen fra 200 nm til 500 nm (avhengig hovedsakelig av den numeriske apertur for det valgte målet, og på den bølgelengde som brukes), og kan lett måles ved hjelp av en kalibreringseksperiment nano- store fluoriserende perler. Thus, kan en pikselstørrelse på 70-150 nm (tre ganger lavere enn den instrument midje) være nok. Imidlertid kan størrelsen pixel være tilpasset til systemet som studeres tar hensyn til en enkel regel: senke pikselstørrelse, høyere nøyaktighet i beskrivelsen av korrelasjonsfunksjonen. Videre har minimumsstørrelsen på bildet for å bli kjøpt til å være minst tre ganger større enn den maksimale forskyvning av interesse (pluss instrumental midje). Dette er nødvendig for å oppnå en god konvergens av tilpasningsalgoritme, og en statistisk signifikant utvalg av molekylære forskyvninger. Som et eksempel, for å studere midlere molekyl forskyvninger som er mindre enn noen få hundre nanometer (f.eks, 200 nm) er et bilde av størrelse noen få mikrometer nok. Videre det totale antall piksler (tar konstant pikselstørrelsen) virkninger på kvaliteten på korrelasjonsfunksjonen. Faktisk tillater et større bilde gjennomsnitt mer informasjon i korrelasjonsfunksjonen, selv om i det bekostning av tiden Resolution. Når det gjelder kamerabasert system som brukes her, vær oppmerksom på at den fysiske størrelsen på pixel på brikken er fast. Følgelig reduserte størrelsen piksel senker signal i hvert bildepunkt (som er avhengig av kvadratet av størrelsen piksel), reduserer synsfeltet, og krever høyere forstørrelse strøm. På den annen side, i et skannesystem, hvor observasjonsområdet er fast, reduserte størrelsen piksel vanligvis resulterer i et økt antall piksler på bekostning av tiden oppløsning.
Noen detaljer om detektoren brukes må diskuteres. I motsetning til enkeltfotondetektorer, EMCCD systemer måler en gjennomsnittsintensitet (digital nivå, DL) som ikke er direkte proporsjonal med det oppsamlede lyset på grunn av tilstedeværelse av en forskyvning. Selv om dette offset er lav i forhold til den dynamiske rekkevidden til kameraet (noen hundre sammenlignet med 2 16 i 16 bit avlesning) og ubetydelig i eksperimenter hvor mange fotoner er samlet, må det tas hensyn tiloppnå en riktig normalisert korrelasjonsfunksjon. Også forskyvningen kan brukes som en referanse i forhold med lite lys for å identifisere den mengde av signalet oppsamlet. Videre, for å anslå den gjennomsnittlige mengden av fotoner som samles under oppkjøpet, gjennomsnittlig digital nivå forbundet til hver innsamlet foton må måles. Denne mengden kan hentes ved å utsette kameraet til en svært lav lysintensitet (f.eks bakgrunnen lyset i rommet); faktisk, i dette tilfellet, kan vi med rimelighet anta at bare enkle fotonene komme til kamera, det vil si den målte intensitet kan være relatert til null eller ett foton bare.
Til slutt, la oss kommentere hvordan noen alternative oppkjøps systemer (dvs. ulike mikros oppsett) kan brukes til å utføre de presenterte målinger. Først av alt, kan den "W'factor i ligning 2 (som representerer autokorrelasjon av den instrument PSF) tilpasses til tHan bestemt innsamlingssystem som brukes for å tilpasse de eksperimentelle korrelasjonsfunksjonen. Som tidligere vist 34, er en enkel sak kjøpet whit en laser scanning mikroskop når skannehastigheten er betydelig høyere enn partikkel dynamikk. I et slikt tilfelle, faktisk bevegelse av partiklene i løpet av anskaffelses tid (dvs. linje-tid) kan betraktes som ubetydelig, og korrelasjonsfunksjon er godt tilnærmet ved en gaussisk funksjon. I sammenheng med de nye bildeteknologi, er en interessant tilnærming basert på muligheten til å produsere svært tynne lyse ark (1-2 mm) gjennom prøven 41. Lyset arket tillater selektiv belysning av et enkelt plan (Single plane mikros belysning, SPIM) i prøven, og kombinert med en kamerabasert tilegnelsessystem, fast optisk snitte i 3D 42. På grunn av disse egenskapene, har SPIM blitt konjugert med FCS 43 og kunne representere en valid verktøy for å forlenge den fremlagte analysen til 3D-miljøer, så som i cytoplasma, eller kjernen av levende celler.
Oppsummering, fra et eksperimentelt synspunkt denne tilnærmingen krever kun tilgang til et mikroskop utstyrt med en rask anskaffelses modul. Protein av interesse kan være merket med fluorescerende protein eller organisk fluoroforen, og dermed gir også for flerfarget imaging. I denne sammenheng ser vi for oss muligheten til å bruke kryss i MSD analyse for å velge sub-populasjoner av molekyler og avsløre interaksjoner og co-diffusjon på live cellemembraner. Til slutt tror vi at denne tilnærmingen kan representere et kraftig verktøy for å studere proteiner og / eller fett inne i en dynamisk partisjonering innen nanodomains på plasmamembranen. I dette tilfellet, den svært variabel størrelse og levetid nanodomains innføre et ekstra nivå av kompleksitet i det virkelige data som ville kreve ytterligere metodiske implementeringer inkludert to-farge imaging, lokaleAnalyse (for eksempel 2D par korrelasjon) og / eller fluorescens anisotropi.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| iXon Ultra 897 | Andor | DU-897U-CS0 | |
| Solis | Andor | ||
| CHO-K1 | ATCC | CCL-61 | |
| ATTO 488 labeled PPE | ATTO-TEC GmbH | AD 488-151 | |
| DOPE | Avanti Polar Lipids, Inc. | 850725 | |
| DOTAP | Avanti Polar Lipids, Inc. | 890890 | |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | |
| DMEM/F-12 | Gibco | 21331 | |
| FBS | Gibco | 10082147 | |
| HEPES | Gibco | 15630-106 | |
| PBS | Gibco | 10010-023 | |
| SimFCS 3.0 | Globals Software | the software can be downloaded here: http://www.lfd.uci.edu/globals/ | |
| DMI6000 with TIRF modulus | Leica | ||
| LAS AF | Leica | ||
| Lipofectamine 2000 | Lipofectamine | 11668019 | |
| Matlab | MathWork | ||
| ImageJ | NIH |
| Name | Company | Catalog Number | Comments |
| C-terminal GFP tagged Tranferrin Receptor | OriGene | RG200980 | |
| Agar | Sigma Aldrich | A5306 | |
| Chloroform | Sigma Aldrich | 528730 | |
| Latex beads, fluorescent yellow-green, 30 nm | Sigma Aldrich | L5155 | |
| SONICA Ultrasonic Cleaners | SOLTEC | ETH S3 | |
| Petri Dishes | Willco | GWSt-3522 | |
| Bio-Format importer for Matlab | http://www.openmicroscopy.org/site/support/bio-formats5/users/matlab/ | ||
| ICS-MatLab Tools | https://www.cellmigration.org/resource/imaging/software/ICSMATLAB_28-02-06.zip | ||
| Simulation by Matlab Tutorial | https://www.cellmigration.org/resource/imaging/icsmatlab/ICSTutorial.html | ||
| Simulation by SimFCS Tutorial | https://www.cellmigration.org/resource/imaging/ppt-pdf/RICS%20Simulations.ppt |
References
- Engelman, D. M. Membranes are more mosaic than fluid. Nature. 438 (7068), 578-580 (2005).
- Vereb, G., et al. yet structured: The cell membrane three decades after the Singer-Nicolson model. Proc. Natl. Acad. Sci. U. S. A. 100 (14), 8053-8058 (1073).
- Ishihara, A., Hou, Y., Jacobson, K. The Thy-1 antigen exhibits rapid lateral diffusion in the plasma membrane of rodent lymphoid cells and fibroblasts. 84 (5), 1290-1293 (1987).
- Axelrod, D., et al. Lateral motion of fluorescently labeled acetylcholine receptors in membranes of developing muscle fibers. Proc. Natl. Acad. Sci. U. S. A. 73 (12), 4594-4598 (1976).
- Jacobson, K., Derzko, Z., Wu, E. S., Hou, Y., Poste, G. Measurement of the lateral mobility of cell surface components in single, living cells by fluorescence recovery after photobleaching. J. Supramol. Struct. 5 (4), 10-1002 (1976).
- Kusumi, A., et al. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annu. Rev. Biophys. Biomol. Struct. 34, 351-378 (2005).
- Kusumi, A., Ike, H., Nakada, C., Murase, K., Fujiwara, T. Single-molecule tracking of membrane molecules: plasma membrane compartmentalization and dynamic assembly of raft-philic signaling molecules. Semin. Immunol. 17 (1), 3-21 (2005).
- Schwille, P., Korlach, J., Webb, W. W. Fluorescence correlation spectroscopy with single-molecule sensitivity on cell and model membranes. Cytometry. 36, 176-182 (1999).
- Gielen, E., et al. Diffusion of sphingomyelin and myelin oligodendrocyte glycoprotein in the membrane of OLN-93 oligodendroglial cells studied by fluorescence correlation spectroscopy. C. R. Biol. 328 (12), 1057-1064 (2005).
- Weiss, M., Hashimoto, H., Nilsson, T. Anomalous protein diffusion in living cells as seen by fluorescence correlation spectroscopy. Biophys. J. 84, 4043-4052 (2003).
- Wawrezinieck, L., Rigneault, H., Marguet, D., Lenne, P. F. Fluorescence correlation spectroscopy diffusion laws to probe the submicron cell membrane organization. Biophys. J. 89 (6), 4029-4042 (2005).
- Lenne, P. F., et al. Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J. 25 (14), 3245-3256 (2006).
- Ries, J., Schwille, P. Studying slow membrane dynamics with continuous wave scanning fluorescence correlation spectroscopy. Biophys. J. 91 (5), 1915-1924 (2006).
- Ruan, Q., Cheng, M. A., Levi, M., Gratton, E., Mantulin, W. W. Spatial-temporal studies of membrane dynamics: scanning fluorescence correlation spectroscopy (SFCS). Biophys. J. 87 (2), 1260-1267 (2004).
- Berland, K. M., So, P. T., Chen, Y., Mantulin, W. W., Gratton, E. Scanning two-photon fluctuation correlation spectroscopy: particle counting measurements for detection of molecular aggregation. Biophys. J. 71, 410-420 (1996).
- Heinemann, F., Betaneli, V., Thomas, F. A., Schwille, P. Quantifying lipid diffusion by fluorescence correlation spectroscopy: a critical treatise. Langmuir. 28 (37), 13395-13404 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Capturing directed molecular motion in the nuclear pore complex of live cells. Proc. Natl. Acad. Sci. U. S. A. 109 (25), 9863-9868 (2012).
- Sanchez, S. A., Tricerri, M. A., Gratton, E. Laurdan generalized polarization fluctuations measures membrane packing micro-heterogeneity in vivo. Proc. Natl. Acad. Sci. U. S. A. 109 (19), 7314-7319 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Fluorescence correlation spectroscopy of intact nuclear pore complexes. Biophys. J. 101 (4), 27-29 (2012).
- Di Rienzo, C., et al. Unveiling LOX-1 receptor interplay with nanotopography: mechanotransduction and atherosclerosis onset. Sci. Rep. 3, 10-1038 (2013).
- Unruh, J. R., Gratton, E. Analysis of molecular concentration and brightness from fluorescence fluctuation data with an electron multiplied CCD camera. Biophys. J. 95 (11), 5385-5398 (2008).
- Kannan, B., et al. Electron multiplying charge-coupled device camera based fluorescence correlation spectroscopy. Anal. Chem. 78 (10), 3444-3451 (2006).
- Jones, S. A., Shim, S. H., He, J., Fast Zhuang, X. three-dimensional super-resolution imaging of live cells. Nat. Methods. 8 (6), 499-508 (2011).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy. 3 (10), 793-795 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Hess, S. T., Girirajan, T. P., Mason, M. D. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys. J. 91 (11), 4258-4272 (2006).
- Manley, S., et al. High-density mapping of single-molecule trajectories with photoactivated localization microscopy. Nat. Methods. 5 (2), 155-157 (2008).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Klar, T. A., Hell, S. W. Subdiffraction resolution in far-field fluorescence microscopy. Opt. Lett. 24 (14), 954-956 (1999).
- Eggeling, C., et al. Direct observation of the nanoscale dynamics of membrane lipids in a living cell. Nature. 457 (7233), 1159-1162 (2009).
- Hedde, P. N., et al. Stimulated emission depletion-based raster image correlation spectroscopy reveals biomolecular dynamics in live cells. Nat. Commun. 4, Forthcoming.
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophys. J. 88 (5), 3601-3614 (2005).
- Brown, C. M., et al. Probing the integrin-actin linkage using high-resolution protein velocity mapping. J. Cell Sci. 119, 5204-5214 (2006).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proc. Natl. Acad. Sci. U. S. A. 110 (30), 12307-12312 (2013).
- Mueller, V., et al. STED nanoscopy reveals molecular details of cholesterol- and cytoskeleton-modulated lipid interactions in living cells. Biophys. J. 101 (7), 1651-1660 (2011).
- Kleusch, C., Hersch, N., Hoffmann, B., Merkel, R., Csiszar, A. Fluorescent lipids: functional parts of fusogenic liposomes and tools for cell membrane labeling and visualization. Molecules. 17 (1), 1055-1073 (2012).
- Ries, J., Chiantia, S., Schwille, P. Accurate determination of membrane dynamics with line-scan FCS. Biophys. J. 96 (5), 1999-2008 (2009).
- Kolin, D. L., Wiseman, P. W. Advances in image correlation spectroscopy: measuring number densities, aggregation states, and dynamics of fluorescently labeled macromolecules in cells. Cell Biochem. Biophys. 49 (3), 141-164 (2007).
- Digman, M. A., et al. Measuring fast dynamics in solutions and cells with a laser scanning microscope. Biophys. J. 89 (2), 1317-1327 (2005).
- Ritchie, K., et al. Detection of non-Brownian diffusion in the cell membrane in single molecule tracking. Biophys. J. 88 (3), 2266-2277 (2005).
- Voie, A. H., Burns, D. H., Spelman, F. A. Orthogonal-plane fluorescence optical sectioning: three-dimensional imaging of macroscopic biological specimens. J. Microsc. 170, 229-236 (1993).
- Huisken, J., Swoger, J., Del Bene,, Wittbrodt, F., J,, Stelzer, E. H. Optical sectioning deep inside live embryos by selective plane illumination microscopy. Science. 305 (5686), 1007-1009 (2004).
- Wohland, T., Shi, X., Sankaran, J., Stelzer, E. H. Single plane illumination fluorescence correlation spectroscopy (SPIM-FCS) probes inhomogeneous three-dimensional environments. Opt. Express. 18 (10), 10627-10641 (2010).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}