

Abstract
它已成为越来越明显,空间分布和膜元件,如脂质和蛋白质的运动是关键因素在许多细胞功能的调节。然而,由于快速的动力学和所涉及的微小结构,非常高的时空分辨率,需要捕捉分子的真实行为。这里,我们提出的实验方案用于研究活细胞中以高时空分辨率荧光标记的质膜蛋白质和脂质的动态。值得注意的是,这种方法并不需要跟踪每个分子,但它计算使用的所有分子在膜中的一个给定区域的人口行为。其出发点是在膜上某一区域的快速成像。之后,一个完整的时空自相关函数计算在增加的时间延迟,例如每2,3中,n次重复关联采集的图像。因此能够证明该宽度的在增加的时间延迟作为粒子运动的函数,由于扩散空间自相关函数的增加的峰值。因此,串联的自相关函数的拟合使得能够提取实际蛋白质意味着从成像(IMSD)方位移,这里提出的表观扩散系数比平均位移的形式。这就产生了单分子与纳米精度的平均动态的定量看法。通过使用转铁蛋白受体(TFR)和一个ATTO488的GFP标记的变体标记的1 -棕榈酰基-2 -羟基- SN -glycero-3-磷酸乙醇胺(PPE),可以观察到的蛋白质和脂质扩散的时空调控上微米尺寸的膜的区域中的微至毫秒的时间范围内。
Introduction
从原来的“流体镶嵌”模型歌手和尼科尔森开始,细胞质膜的画面不断在过去几十年更新,以包括细胞骨架和脂质域1,2的新兴角色。
第一观察由荧光漂白恢复(FRAP)揭开该膜蛋白的显著分数是不动的3-5后得到。这些开创性的研究,虽然非常丰富,受到FRAP设置在空间(微米)相对较差的分辨率和时间(秒)。此外,作为一个统计平均测量,FRAP缺乏使单分子行为的信息。
在这种情况下,有可能以特别标示具有非常明亮的标记(允许在扩散过程中的一个分子在一个时间的研究)的单个分子已经非常成功。特别是,通过推动单粒子追踪(SPT)的方法来微秒的时间尺度,久住,等脂质和蛋白质动力学的未知功能,极大地促进了表彰肌动蛋白为基础的膜骨架的作用在细胞膜的生理6 人获得了进入的时间分辨率,7。这些发现产生了所谓的“尖桩和栅栏”的模式,在其中的脂质和蛋白质的扩散是通过基于肌动蛋白骨架的调节。然而,为了能够访问大量的由SPT许多实验的问题所提供的信息必须得到解决。具体地讲,标记过程通常是通过许多步骤,如标记物种进入该系统的生产,纯化和介绍组成。此外,大的标签,如量子点,金属纳米颗粒,通常需要达到亚毫秒级的时间尺度与目标分子的标记的交联无法避免在许多情况下。最后,许多轨迹已被记录,以符合统计标准和伴随低密度标签的需要,以允许跟踪。
相比SPT,荧光相关光谱(FCS),克服许多这些缺点,是一种非常有前途的方法来研究分子动力学。事实上,FCS的效果很好也有暗淡而密的标签,使研究荧光蛋白标记的分子动力学瞬时转染细胞。此外,它允许达到较高的数据在有限的时间。最后,尽管标签的“高”密度FCS提供单个分子的信息。感谢所有这些特性,FCS是一个非常简单的方法,并已被广泛应用于研究脂类和蛋白质动力学无论是在模型膜和活细胞8-10。许多不同的方法已经被提出以提高FCS的揭示分子扩散的细节的能力。例如,它是sh的自己通过对不同大小的观察区域进行的FCS可以定义分子运动11,12的“FCS扩散法”的启发隐藏的功能。除了 被大小不一,重点领域也重复13,移动沿线14-20或共轭快速相机21,22空间。使用这些“时空”的相关办法,几个膜元件相关的生物学参数上都细胞膜的模型与实际的生物学者,由此制得的洞察膜空间组织进行了定量描述。
然而,在所有的FRAP和FCS应用到目前为止描述的焦点区域的大小代表了在空间分辨率的限制,无法克服。几个超分辨率成像方法近来已经开发绕过此限制。有些是基于定位的精度,如随机光学重建显微镜(STORM)<SUP> 23,24,光活化定位显微镜(PALM)25,荧光PALM(FPALM)26,和单粒子跟踪PALM(sptPALM)27:比较大的值,在每个快照需要的光子,但是,限制的时间分辨率这些方法中的至少几毫秒,从而妨碍了它们在体内的适用性。
与此相反,一个有前途的替代超分辨率成像已被打开的空间调制与受激发射损耗的方法(STED或可逆饱和光荧光跃迁(RESOLFT))28,29的荧光发射。这些方法结合起来观察体积的成形以及低于衍射极限与使用快速扫描显微镜和检测系统的可能性。在用荧光波动分析组合,STED显微镜允许直接探测脂质和p的纳米尺度的时空动力学roteins活细胞膜30,31。
相同的物理量的受激发射损耗系显微镜可以通过改性时空图像相关光谱(STICS 32,33)方法,该方法适合于荧光标记的膜蛋白和/或脂 类中的活细胞的动力学的研究得到而由商业显微镜。实验方案这里介绍的是由几个步骤组成。第一个要求在膜感兴趣区域的快速成像。然后,将得到的图像的堆栈用来计算平均值的空间 - 时间相关性的函数。通过拟合一系列的相关函数,该分子“扩散法”,可直接从成像获得的表观扩散系数(D 应用程序 )的形式- 比 -平均位移曲线图。该图的关键取决于由分子探索的环境,并允许直接识别的实际扩散模式的感兴趣的脂质/蛋白质。
更详细地,如先前所示图34中,获取的图像序列的时空自相关函数的关键取决于移动中所收集的图像序列的分子动力学(请注意,相同的推理也可以从行采集应用凡在空间只是一个维度考虑)。特别地,我们定义的相关函数为:
 (1)
(1)
其中 表示测得的荧光强度中的位置x,y和时间t,
表示测得的荧光强度中的位置x,y和时间t, ![]() 和
和![]() 表示在x的距离和分别为Y方向,
表示在x的距离和分别为Y方向, ![]() 表示的时间差,并
表示的时间差,并![]() 表示平均。这个函数可以表示为:
表示平均。这个函数可以表示为:
 (2)
(2)
其中“N”表示在该观察区域分子的平均数目, ![]() 表示空间中的卷积运算,并
表示空间中的卷积运算,并 代表器乐腰部的自相关。后者可以被解释为对如何将单个发射器的光子被摊开在空间的量度,由于光/记录设置程序(所谓的点扩散函数,PSF,基因涨势以及由高斯函数近似)。最后,
代表器乐腰部的自相关。后者可以被解释为对如何将单个发射器的光子被摊开在空间的量度,由于光/记录设置程序(所谓的点扩散函数,PSF,基因涨势以及由高斯函数近似)。最后,  代表的概率找到一个粒子的距离
代表的概率找到一个粒子的距离![]() 和
和![]() 延时后,
延时后, ![]() 。如果我们考虑扩散动力学,粒子随机移动在各个方向和净通量是不存在的,这个功能也很好用高斯函数,其中的差异,可以认定为运动粒子的均方位移(MSD)的近似。因此,相关函数的腰部(也称为
。如果我们考虑扩散动力学,粒子随机移动在各个方向和净通量是不存在的,这个功能也很好用高斯函数,其中的差异,可以认定为运动粒子的均方位移(MSD)的近似。因此,相关函数的腰部(也称为![]() ),可以被定义为颗粒的MSD的总和和器乐腰部,并且可以通过高斯拟合来测定婷的相关函数为每个时间延迟。在测量电流I MSD可以用来计算移动分子的表观扩散系数
),可以被定义为颗粒的MSD的总和和器乐腰部,并且可以通过高斯拟合来测定婷的相关函数为每个时间延迟。在测量电流I MSD可以用来计算移动分子的表观扩散系数![]() 和平均位移
和平均位移![]() 如:
如:
 (3)
(3)
 (4)
(4)
很少考虑对实验装置可以用来引导读者在以下几节。为了选择性地激发荧光团上的活细胞,我们将使用全内反射(TIR)的照明,使用市售的TIR荧光的基膜(TIRF)显微镜(细节可在材料部分中找到)。此外,为了收集日ê荧光,我们将使用高倍率物镜(100X NA 1.47,高数值孔径需要全内反射荧光照明)和EMCCD摄像头(像素的物理尺寸上的芯片16微米)。以达到100nm的像素大小,我们应用1.6X的附加放大率透镜。如下面讨论的,将需要低于1毫秒的时间分辨率来适当地描述的快速膜脂质的动力学低于100nm。为了达到这个时间分辨率,我们需要选择的感兴趣区域(ROI)的区域比相机(512×512)的整个芯片更小。在这种方式中,相机将读取线增加时间分辨率的数目可以减少。然而,在此读出机制的帧时间将通过从曝光的电荷转移到读出芯片的照相机上需要与通常以毫秒为512×512像素的EMCCD的顺序的时间限制。要打败这个限制,一个新兴的技术可以转移的投资回报率,仅行而不是整个框架,W第i个实际有效减少暴露的芯片尺寸(称为我们的EMCCD七分传感器模式)。对于这种配置是有效的,必须将ROI的外片覆盖的一对狭缝安装在光路中的。由于这种设置的时间分辨率降低到10 -4秒就可以实现。但是,请注意,这种方法可以与许多不同的实验装置,为“讨论”一节中的说明。
该方法的演示将在活细胞中提供,通过使用这两个标记的一个ATTO488 1 -棕榈酰基-2 -羟基- SN -glycero-3-磷酸乙醇胺(ATTO488-PPE)和转铁蛋白受体的GFP-标记的变体(GFP- TFR)。在ATTO488-PPE的情况下,这种方法可以成功地回收了几乎常数D 的应用程序作为平均位移表示较自由扩散的功能,如先前报道30,35。相比之下,TFR-GFP呈减少ð
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1,系统校准
- 点扩散函数(PSF)的校准
- 稀释10微升的30纳米荧光珠溶液(约5μm)在90微升蒸馏水中,然后超声处理20分钟,该溶液中。切成正方形(1 1cm×1cm的)一块琼脂糖凝胶(3%)和存款10微升在凝胶顶部的溶液中。推翻了一块凝胶上2厘米的培养皿底部的玻璃和挤压玻璃上的下降。
- 打开采集设置,把样品中的持有人,将相机的曝光和EMgain(100毫秒和1000都不错的参数,但优化根据系统),并等待相机冷却下来。
- 将相机的曝光为100毫秒,相机EMgain到1000,采集模式帧传输,100重复和自动保存设置。
- 使用目镜和透射的光聚焦在凝胶上的边界,然后移动目标的凝胶的中心,调整焦距和站RT激光校准程序(在拉斯维加斯的AF,选择'全内反射荧光设置“,然后按自动调整过程)。
- 找到的视场与分离的单斑点,准确地集中于较亮的点(即通常代表珠子聚集体)作为参照,取得100帧,以便获得多个单点重复步骤5-6次。
- 导入所获得的一系列的数据处理程序,并在时间平均化叠层( 图1A),并选择单个孤立的珠子。小心选择最小的,以避免粒子的聚集体。
- 适合使用命令“gaussfit”所选择的光强分布(单珠信息的一例示于图1B)用高斯函数(在ICS-Matlab的工具中的材料在Matlab)。通过检查得到的残差检验的拟合优度(拟合高斯分布与相应的残差的一个例子,提出íQ 图1B)。
- 摄像机标定
- 打开相机电源,等待摄像机冷却下来。将摄像头采集的设置( 即 ,对于所使用的摄像头,我们将曝光到0.5毫秒,相机EMgain到1000,采集模式,以裁切模式,投资回报率的大小为32×128,万家重复),并开始采集摄像头背景信号。
- 导入收购框架系列的数据处理程序。计算并检查在每个像素的平均强度,以确认该照相机的背景是在芯片的选择的区域大致平坦。在裁切模式,除去第一和最后几水平线(3至10根据ROI的大小)对每一帧,因为相机的背景通常是偏向于边界线。
- 创建值的直方图(也定义了电子水准仪,DL)中获得的图像栈(在Matlab中使用命令“HIST”),并绘制对数产生的频率(在Matlab使用semilogy命令)。 DL分配的相机背景的一个例子示于图2。
注:如果相机运作良好,剧情会显示约高斯峰(对数尺度的抛物线轮廓),代表相关零光子之后的指数衰减(以对数刻度负斜率的线值的分布),表示相关联的1光子( 图2)的值的分布。特别是,中心和高斯函数的方差表示相机的偏移和误差,分别,而指数部分的衰减常数表示由相机对每个单光子分配的DL的估计。在Matlab中使用脚本的一节“CalibrateCamera”的配套教材。 - 重复上述操作所有选定相机EMGain和增益。
2,标记细胞的制备
- 以制备所需的脂质掺入36的脂质体,溶解分别为1mg DOPE(1,2-dioleoyl- SN -glycero-3-磷酸乙醇胺),1毫克DOTAP(1,2 -二油酰-3 -三甲基铵-丙烷)的和于1ml氯仿中1毫克PPE-ATTO488的。混合在一起加入0.5ml DOPE溶液,0.5毫升DOTAP溶液,并加入25μl的PPE-ATTO488溶液并在真空下干燥24小时。加入0.5毫升HEPES缓冲液为20mM的,涡旋15分钟,超声处理15分钟,在40℃下。
- 以制备的细胞,用PBS洗3次汇合的CHO-K1(中国仓鼠卵巢)的P100菜,在培养箱中加入1 ml胰蛋白酶和商店的5分钟。暂停分离的细胞中加入9毫升DMEM / F12培养基补充有FBS的10%,种子150微升的细胞溶液中含有800微升的同一种培养基的培养皿中。
- 商店在培养箱中培养24小时,在37℃和5%CO 2。脂质结合,取代500细胞培养基微升无血清的培养基中; 30分钟后,加入2微升的脂质体溶液; 15分钟后洗涤用PSB并添加新的DMEM / F12培养基以进行成像。
- 对于转染,根据使用转铁蛋白-GFP质粒,并存储24小时在恒温箱成像之前的Lipofectamine协议(制造商的说明)转染细胞。
3,数据采集
- 安装准备
- 为了温控显微镜,实验前24小时开机的孵化器。
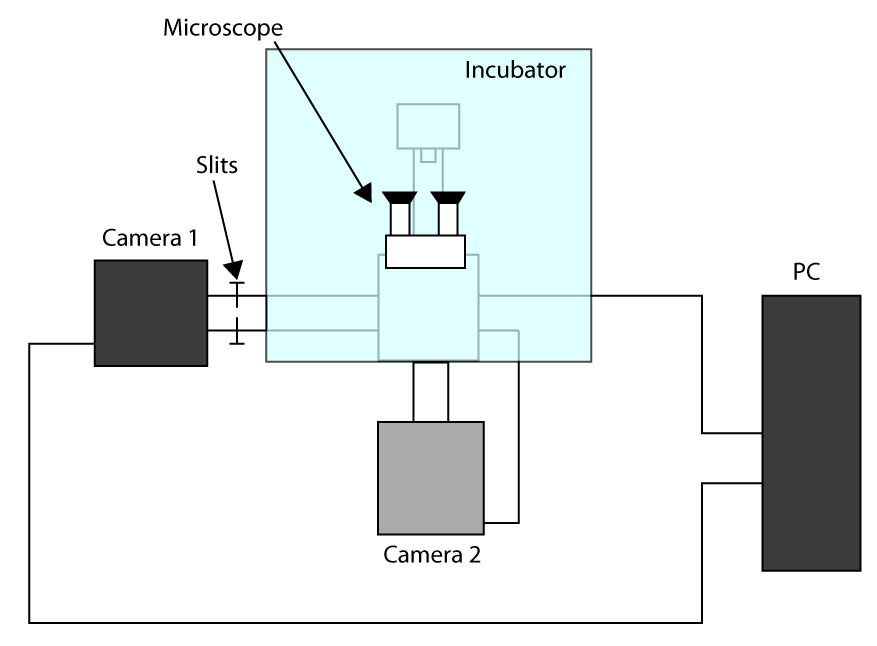
- 为了应用最快达到的采集时间,在裁剪传感器模式工作(见介绍),并使用第一相机的成像(相机1)和第二相机来选择小区(相机2)。安装配置的方案呈现在补充图S1。然后,对准两个相机开启显微镜和等待相机冷却下来。
- 将两款相机的参数,透射光成像( 即
- 将样品在支架和重点用目镜,送光相机1,轻轻推了开衩让光只用于细胞成像(这里是32×32像素的投资回报率)的投资回报率。
- 移动一个单元格选定区域和发送光相机2,然后绘制在控制摄像头2为了有一个参考该软件的投资回报率。
- 成像( 图3A)
- 首先,根据您的设置的程序对齐全内反射荧光激光。在我们的设置中,选择“全内反射荧光设置”,并开始自动调整过程。当激光对准设置的穿透深度为70nm(约70°)。
- 设置曝光时间为70毫秒,EMGain 100两个摄像机1和摄像机2;然后,选择用相机1的单元格,然后发送光相机2,准确地聚焦在细胞膜上。坐落在加利福尼亚州的最低曝光MERA 2 1000 EMGain,裁剪传感器模式,10 5的重复和设置自动保存为配合文件(灵活图像传输系统,一种格式,可以很容易地进行管理)。
- 开始采集记录的图像系列。释放增益和裁剪方式,以使温度稳定取得了新的电池前,那么为了获得8-10细胞重复最后两个步骤。
从影像学4,计算的均方位移(I MSD)
注意:下面的协议可以被直接应用到原始数据。同时,在整个协议都有效在Matlab和在SimFCS模拟数据采集。链接到相应的教程可以在“材料”部分中找到。
- 计算用Matlab
- 导入使用ImportImageSeries脚本获取的系列Matlab的。使用共同计算各图像的平均亮度在时间mmand意味着在第一个2的尺寸和使用小区看到生成的载体。
- 若漂白的10%以上存在时,丢弃该系列或删除它们的第一部分。如果是较低的,尝试纠正对相关函数的影响中减去每个图像的平均强度,如37之前所示。
- 通过使用平均值在第三维度计算每个像素的平均强度和见得到的图像。
注意:要求特别注意以避免人为的相关性。事实上,如前面对于类似的技术38,单元格边框以及聚焦囊泡所示可以引入很强的相关性。如果平均图像的检查结果显示单元格边框或失焦小泡,尽量排除参与,否则丢弃该收购的区域。要纠正这种不动结构的影响,从每个像素39减去平均时间上的强度。 - 计算吨他的时空相关性(G(ξ,χ,τ))的使用功能CalculateSTICScorrfunc。删除G(ξ,χ,0),因为由于在低光环境的散粒噪声的相关性主导G(0,0,0);由于检测器的相关主导着G(±1,0,0),和粒子运动过程中的曝光时间可变形G(ξ,χ,τ)为τ= 0,通过增加测量的腰部(该效果消失为τ > 0)34。
- 平均g用对数时间斌用“LogBinStack”功能的辅助材料以减少噪声,然后(ξ,χ,τ> 0)满足所产生的G(ξ,χ,τ)使用功能“gaussfit”在材料的ICS与Matlab工具恢复的I MSD(结果数组的第二列)。
- 积所得的腰部σ(τ)2(ⅰMSD)作为时间的函数。如果数据过于嘈杂,设法增加acqu数IRED帧,增加激光功率,平均多个G(ξ,χ,τ)在一起。
- 计算由SimFCS
- 与使用ImageJ BioFormat进口商插件打开文件获取并保存后天系列为TIFF序列。
- 打开SimFCS并选择RICS工具,然后选择文件>导入多张图片(补充图S2)。
- 选择飞度,插入正确的采集参数,并关闭窗口合身(补充图S3)。
- 选择显示>平均强度> CH1和验证漂白的存在(补充图S4)。
- 若漂白的10%以上是本丢弃的系列,或者如果它是可能的负载再次在图像序列中除去该系列的第一部分。
- 如果漂白是低于10%,选择工具>我的MSD>设置参数,选中“使用移动平均线”,在左边一个在ROI面板中设置赭帧的移动平均关注的对应时间比特征扩散时间增加(为粒子在1微米2秒移动-1 10秒的时间是一个很好的移动平均)
- 选择工具> IMSD>计算ìMSD(补充图S5),并配合和记事簿导出的I MSD(补充图六 )。
从I MSD 5计算扩散法
- 适合的前几个点来推断的截距(σ0 2)(5分通常是足够的,但更多的点可以安装如果它们表现出非线性行为),该值与先前测量的PSF 2比较。如果它们是可比较的,孤立的荧光动力学正紧随其后。与此相反,如果σ0 2 >> PSF 2试图获取更快,以确保没有任何隐藏的动力是目前的34。
- 计算表观扩散系数(D 应用程序 ),并使用等式3和4(参见引言)的平均位移(R)。
- D地块的应用程序为R的函数来获得一个扩散定律什么是衡量基于FCS的12( 图3D)点的变化相媲美。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
为了校准的仪器腰部,单个荧光纳米珠的图象可以作为在协议步骤1.1中描述为量度。这些珠的典型的荧光图像示于图1。强度分布由二维高斯函数拟合还给良好的残差,并允许测量仪器腰部在270nm处。这个值是在与由瑞利方程估计的预期衍射极限相吻合。该校准是没有必要的粒子动力学的测量,但它需要测量的表观粒度。
相机背景的典型频率分布示于图2。在约180 DL的峰值是由于照相机响应于没有光子,并且它代表模拟数字(AD)转换器的贡献。这种贡献可以近似为高斯分布来估计偏移和方差由信号记录引入。高于200的DL的数字电平分布成为指数(线性对数标度),表示平均的相机响应单个光子。装修这部分与指数分布可以分配给每个单光子的平均DL的测量。较高的是分配给每个光子的平均DL和AD转换误差之间的比率,下部将在计算出的相关函数的噪声。另外,平均单光子响应允许摄像机动态范围的估计。
完整的实验步骤的示意图总结在图3和Atto488-PPE插入的图片进入膜被表示在图4A中。为CHO细胞标记Atto488-PPE的基底膜的代表全内反射荧光图像显示在图4B。有几个亮点,可能是由于李在细胞外存在堆叠在玻璃posomes。它们可以通过在膜部选择的ROI大多均匀的荧光被丢弃( 即 ,蜂窝质膜)。如预期所测量的扩散法( 图4C),用于此脂质是平的,指示先前由STED-FCS测量30,35所示的较自由扩散。值得一提的是,所有的所示的位移值是低于衍射极限,清楚地表明这种方法的能力的超解析远低于衍射极限的上下均分子量位移到几十纳米。
甲图式转铁蛋白-GFP二聚体插入到膜被表示在图5A中 。许多研究表明,该受体的胞质尾相互作用与膜骨架,这反过来又用作栅栏对受体的流动性12,40。 CHO细胞表达转铁蛋白受体-GFP的代表全内反射荧光图像presented 图5B中。低荧光强度的细胞应该是首选,因为该膜是更接近天然状态和相关的过度表达被最小化伪影的可能性。此外,应避免使用的小区的中心部分,由于离焦的荧光的影响(从细胞质中,例如)可能存在。如预期所测量的扩散法( 图5C)的转铁蛋白-GFP显示了低于100nm的第一平坦的行为,以约0.7微米2秒-1的平均ð 应用程序 ,随后在表观扩散系数随之迅速下降,范围在0.2微米2秒-1(该值通常通过测量衍射限制的FCS 12)。这一结果表明,我们的方法可以很容易地测量绿色荧光蛋白标记的蛋白质的平均位移与几十纳米的分辨率。此外,该空间刻度D 应用程序开始时的降低套的特点由细胞膜骨架蛋白部分封闭空间尺度约为120纳米,符合先前估计6。

图1。校准点扩散函数。 (A)中的分离的珠子和珠子聚集体的伪彩色图象。分离的胎圈的强度轮廓(B)的三维图显示了良好定义的高斯分布(C)由高斯函数中的强度分布的拟合(上图)与相应的残差(下图)。拟合分布和测得的强度分布之间良好的一致性也是一个证明,器乐PSF可以用高斯函数来近似。 请点击这里为vi EW这个图的放大版本。

图2:校准相机响应单光子。图中显示的数字水准仪(DL)分布于相机的背景在一个32×128的投资回报率,曝光0.5毫秒,在裁剪传感器模式。在约180 DL的峰值代表了相机响应于没有光子。具体地讲,它表示的模拟数字(AD)转换器的贡献,并且可以由一个高斯函数来近似估计的偏移和由信号记录引入的方差。高于200的DL数字电平的分布变得指数和代表平均照相机响应单个光子。这些参数的测量允许推定的采集期间被记录的光子密度。S / ftp_upload / 51994 / 51994fig2highres.jpg“目标=”_ blank将“>请点击这里查看该图的放大版本。

图3图式的方法。 (A)的大视场成像通过EMCCD相机被施加到达到亚毫秒级的分辨率,而全内反射荧光显微术被利用来提供质膜的精确光学切片(B)的图像所得到的叠层的自相关,以计算平均空间-temporal相关函数。此相关函数由一个高斯函数很好地近似(参见导言),它伸展在时间根据粒子的位移(C)因此,为了量化该相关函数的扩展,由于分子的位移,用高斯拟合伊恩功能被执行。这允许直接从成像测量分子“扩散法”,在表观扩散系数比平均位移曲线的形式(D)由于该曲线,分子扩散模式可以直接与不需要的解释性模型或假设所识别的空间组织的膜。事实上,可自由扩散的分子将显示一个恒定表观扩散系数作为其流动性并不依赖于测量的空间尺度。相比之下,局部限制的分子将显示一个相当恒定表观扩散系数对位移超过限制大小较小,则减小的扩散率的空间尺度大于限制大小。因此,在表观扩散系数的降低的外观可以被解释为瞬态约束的指纹,而相关的空间尺度可用于估计该限制的空间延伸。 .jove.com /文件/ ftp_upload / 51994 / 51994fig3highres.jpg“目标=”_ blank将“>请点击这里查看该图的放大版本。

图4 ATTO488-PPE扩散法活细胞膜(一)ATTO488-PPE插入细胞膜的示意图(B)的CHO基底膜标记ATTO488-PPE全内反射荧光图像:一个投资回报率(红盒)被选中在小区的大多是制服的一部分,避免了单元格边框和高的荧光斑点。(三)在选定的ROI测量扩散定律表明平坦的行为,确认一个自由扩散模型这个组件。 请点击这里查看大图这个数字。

图5 TFR-GFP的扩散定律活细胞膜(一)TFR-GFP插入细胞膜的示意图。受体的胞质尾相互作用与膜骨架,充当护栏的流动性受体(B。 )CHO表达转铁蛋白受体-GFP的TIRF形象:ROI选择喜欢低表 达的细胞,以避免伪影是由于过表达(C)的转铁蛋白受体(黑点的扩散法),与PPE(灰线,取自图4)。显示部分密闭空间扩散,其中第一平坦部分后面是D中的应用减少的典型行为。 请点击这里查看该网络的放大版本。gure。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
单粒子追踪(SPT)表示的研究分子动力学中最常见的策略之一,它具有测量粒子轨迹的显着优点。这又允许探测在一个复杂的系统,即使少量标记粒子的行为。然而,为了达到这个优点SPT通常需要将探头和非常明亮的标签的密度低。具体地讲,以获得高的时间分辨率(微秒范围)无机探针通常需要( 例如,量子点或金属纳米粒子):在这种情况下的产量,标签和插入一个复杂的过程进系统 是必要的。相较于SPT本发明的方法显示了一些关键的优势。首先,这种方法可以用来在缀合有荧光蛋白。因此,相较于SPT,更高的时间分辨率,实现得益于光子的较 低量(相同的标签上)所需的34。更详细地说,这个属性允许推颞甲阶酚醛树脂低于10 ution - 3秒还使用时可编码的荧光蛋白质,并且这种时间尺度给出了独家获得的膜组成的纳米级动态。最后,值得一提的是,分子扩散法通过分析全时空相关函数,而无需跟踪每个分子中描述。
以受激发射损耗为基础的FCS的比较也很有趣。在STED-FCS测量分子用于降低观测卷的平均传输时间是由荧光信号的时间相关性进行测量。这允许获得的分子动力学也低于衍射极限的局部测量。在所提出的方法,扩散法测量为所有颗粒的平均运动中选定的投资回报率,由标准,衍射限制的,观察体积手段观察。然而,报道的结果表明,该方法不被衍射限制,而是仅由第E,可得时间分辨率。事实上,虽然受衍射限制采集用于检测波动(类似于什么其他超分辨率技术,如PALM和STORM完成),分子位移远低于衍射极限可以是(直接地)计算,如已通过使用STICS测量分子表现出流动32。此外,与STED-FCS,这种方法可以很容易地应用于广泛的商业和现有显微镜的设置,如光栅扫描显微镜或宽视场照相机的基于显微镜。这是值得一提的是分子扩散规律STED-FCS的测量严格要求的仪器腰围尺寸的荧光相关的校准。相反,在测量呈现此处不需要系统校准(仅适用于粒径的估计需要的话)。
在粒子位移的测量通过该方法去实际分辨率暂时搁置关于如何准确地,我们可以测量相关函数。因此,它是不具有固有的衍射限制,类似于向SPT的情况下的分辨率取决于如何准确的粒子“图像”进行测定。以测量在不到1分钟的显著相关的建议的实验中,一些光子(通常低于10的光子),用于在每帧中的每个颗粒都不够。事实上,所有观察到的粒子的贡献被一起平均的相关函数进行计算时,即使颗粒不分离。这个属性是固有的波动相关性的方法,并允许使用暗淡的,致密的标签,如转染的活细胞中的荧光蛋白。
考虑到这一点看来清楚,最小的可测位移取决于颗粒的扩散和在成像设置的时间分辨率。作为一个例子,请考虑细胞膜上的分子的扩散,其中最大测量扩散系数为蛋白质或脂类是大约5微米2秒-1。在这些条件下,我们需要的大约10 -4秒的时间分辨率捕捉的50nm的平均位移。这一次的分辨率可以通过沿单一线或快速的EMCCD相机,在时间分辨率相一致的曝光时间,快速扫描显微镜来实现,如显示在这里。
这种方法的附加基本要求,以准确地描述分子动力学是正确的空间采样。事实上,为了适应该相关函数,我们需要一个空间采样(像素大小)比器乐PSF的腰下部。在大多数商业显微镜(共焦或宽域)中,PSF腰部范围从200纳米至500纳米(主要用于根据所选择的目标的数值孔径和对波长),并且可以通过校准实验使用纳米容易地测量大小的荧光珠。牛逼HUS,70-150纳米(比器乐腰下部的3倍)的像素大小是不够的。然而,像素尺寸可以适合于该系统正在研究考虑一个简单的规则:降低像素尺寸,更高的准确度中的相关函数的说明。此外,图像的可获取的最小尺寸必须在比所关注的最大位移(加器乐腰部)大至少3倍。这是必需的,以便达到拟合算法的良好收敛性和分子位移的统计学显著采样。作为一个例子,来研究均分子量位移大于几百纳米( 例如 ,200 nm)的几微米的图像尺寸是足够小。此外,像素的总数(以恒定的像素大小)的影响的相关函数的质量。事实上,一个更大的图像均允许在相关函数的详细信息,即使在牺牲的时候resolut的离子。关于这里所用的基于相机的系统中,请注意,在芯片上的像素的物理尺寸是固定的。因此,减小了像素尺寸降低了在每个像素中的信号(即取决于像素大小的平方),减小的视场,并且需要较高的放大率。另一方面,在扫描系统中,在观察区域是固定的,减小了像素尺寸通常导致增加的像素数在时间分辨率为代价的。
关于所使用的探测器一些细节还有待讨论。不像单光子探测器,EMCCD系统测量的平均强度(数字电平,DL),其不直接正比于所收集的光由于偏移的存在。即使这个偏移量是比较低的摄像机的动态范围(几百相比,2 16,在16位的读出),并且在许多光子被收集的实验可以忽略不计,它必须被考虑到获得的相关函数的正确正常化。另外,可以用作在低光照条件下的基准,以便确定信号的采集量的偏移量。此外,为了估计在采集过程中所收集的光子的平均量,平均数字电平相关联的每个收集的光子具有被测定。这个数量可以让相机非常低的光强度进行检索( 例如 ,在房间里的背景光);事实上,在这种情况下,我们可以合理地假设,只是单光子到达照相机, 即所测量的强度可以与仅零个或一个光子。
最后,让我们对如何一些备选采集系统( 即 ,不同的显微镜装置),可被用于执行所提出的测量评价。首先,在“W'factor方程2(表示该乐器的PSF的自相关)可以适于吨为了拟合实验相关函数用他特定采集系统。如先前所示34,一个简单的例子是采集白衣的激光扫描显微镜,当扫描速度小于粒子动力学显著更高。在这种情况下,实际上,颗粒在采集时间的移动( 即 ,行时间)可以被认为是可以忽略不计和相关函数是由一个高斯函数很好地近似。在新兴的成像技术的上下文中,一个有趣的方法是基于通过样品41以产生非常薄的光片(1-2微米)的可能性。光片是使试样中的单个平面(单平面照明显微镜,SPIM),并结合了基于照相机的采集系统,快速光学切片的三维42的选择性的照明。由于这些特点,SPIM已成功偶联有FCS 43和可以代表一个瓦利D刀具对所呈现的分析扩展到三维环境中,如在细胞质或活体细胞的细胞核中。
总之,从实验的角度来看这种方法仅需要接入到带有快速采集模块的显微镜。目的蛋白质可以被标记为任何荧光蛋白或有机荧光团,因此允许同时对多色成像。在这方面,我们设想使用交叉ıMSD分析来选择亚群的分子,并揭示于活细胞膜的相互作用和共扩散的可能性。最后,我们认为,这种方法可能是一种有力的工具来研究蛋白质和/或脂质上质膜纳米畴内进行动态分区。在这种情况下,高度可变大小的纳米畴和寿命引入了复杂的真实数据,将需要进一步的方法实现,包括2色成像的附加电平,局部分析( 例如 ,2D对相关性)和/或荧光各向异性。
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| iXon Ultra 897 | Andor | DU-897U-CS0 | |
| Solis | Andor | ||
| CHO-K1 | ATCC | CCL-61 | |
| ATTO 488 labeled PPE | ATTO-TEC GmbH | AD 488-151 | |
| DOPE | Avanti Polar Lipids, Inc. | 850725 | |
| DOTAP | Avanti Polar Lipids, Inc. | 890890 | |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | |
| DMEM/F-12 | Gibco | 21331 | |
| FBS | Gibco | 10082147 | |
| HEPES | Gibco | 15630-106 | |
| PBS | Gibco | 10010-023 | |
| SimFCS 3.0 | Globals Software | the software can be downloaded here: http://www.lfd.uci.edu/globals/ | |
| DMI6000 with TIRF modulus | Leica | ||
| LAS AF | Leica | ||
| Lipofectamine 2000 | Lipofectamine | 11668019 | |
| Matlab | MathWork | ||
| ImageJ | NIH |
| Name | Company | Catalog Number | Comments |
| C-terminal GFP tagged Tranferrin Receptor | OriGene | RG200980 | |
| Agar | Sigma Aldrich | A5306 | |
| Chloroform | Sigma Aldrich | 528730 | |
| Latex beads, fluorescent yellow-green, 30 nm | Sigma Aldrich | L5155 | |
| SONICA Ultrasonic Cleaners | SOLTEC | ETH S3 | |
| Petri Dishes | Willco | GWSt-3522 | |
| Bio-Format importer for Matlab | http://www.openmicroscopy.org/site/support/bio-formats5/users/matlab/ | ||
| ICS-MatLab Tools | https://www.cellmigration.org/resource/imaging/software/ICSMATLAB_28-02-06.zip | ||
| Simulation by Matlab Tutorial | https://www.cellmigration.org/resource/imaging/icsmatlab/ICSTutorial.html | ||
| Simulation by SimFCS Tutorial | https://www.cellmigration.org/resource/imaging/ppt-pdf/RICS%20Simulations.ppt |
References
- Engelman, D. M. Membranes are more mosaic than fluid. Nature. 438 (7068), 578-580 (2005).
- Vereb, G., et al. yet structured: The cell membrane three decades after the Singer-Nicolson model. Proc. Natl. Acad. Sci. U. S. A. 100 (14), 8053-8058 (1073).
- Ishihara, A., Hou, Y., Jacobson, K. The Thy-1 antigen exhibits rapid lateral diffusion in the plasma membrane of rodent lymphoid cells and fibroblasts. 84 (5), 1290-1293 (1987).
- Axelrod, D., et al. Lateral motion of fluorescently labeled acetylcholine receptors in membranes of developing muscle fibers. Proc. Natl. Acad. Sci. U. S. A. 73 (12), 4594-4598 (1976).
- Jacobson, K., Derzko, Z., Wu, E. S., Hou, Y., Poste, G. Measurement of the lateral mobility of cell surface components in single, living cells by fluorescence recovery after photobleaching. J. Supramol. Struct. 5 (4), 10-1002 (1976).
- Kusumi, A., et al. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annu. Rev. Biophys. Biomol. Struct. 34, 351-378 (2005).
- Kusumi, A., Ike, H., Nakada, C., Murase, K., Fujiwara, T. Single-molecule tracking of membrane molecules: plasma membrane compartmentalization and dynamic assembly of raft-philic signaling molecules. Semin. Immunol. 17 (1), 3-21 (2005).
- Schwille, P., Korlach, J., Webb, W. W. Fluorescence correlation spectroscopy with single-molecule sensitivity on cell and model membranes. Cytometry. 36, 176-182 (1999).
- Gielen, E., et al. Diffusion of sphingomyelin and myelin oligodendrocyte glycoprotein in the membrane of OLN-93 oligodendroglial cells studied by fluorescence correlation spectroscopy. C. R. Biol. 328 (12), 1057-1064 (2005).
- Weiss, M., Hashimoto, H., Nilsson, T. Anomalous protein diffusion in living cells as seen by fluorescence correlation spectroscopy. Biophys. J. 84, 4043-4052 (2003).
- Wawrezinieck, L., Rigneault, H., Marguet, D., Lenne, P. F. Fluorescence correlation spectroscopy diffusion laws to probe the submicron cell membrane organization. Biophys. J. 89 (6), 4029-4042 (2005).
- Lenne, P. F., et al. Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J. 25 (14), 3245-3256 (2006).
- Ries, J., Schwille, P. Studying slow membrane dynamics with continuous wave scanning fluorescence correlation spectroscopy. Biophys. J. 91 (5), 1915-1924 (2006).
- Ruan, Q., Cheng, M. A., Levi, M., Gratton, E., Mantulin, W. W. Spatial-temporal studies of membrane dynamics: scanning fluorescence correlation spectroscopy (SFCS). Biophys. J. 87 (2), 1260-1267 (2004).
- Berland, K. M., So, P. T., Chen, Y., Mantulin, W. W., Gratton, E. Scanning two-photon fluctuation correlation spectroscopy: particle counting measurements for detection of molecular aggregation. Biophys. J. 71, 410-420 (1996).
- Heinemann, F., Betaneli, V., Thomas, F. A., Schwille, P. Quantifying lipid diffusion by fluorescence correlation spectroscopy: a critical treatise. Langmuir. 28 (37), 13395-13404 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Capturing directed molecular motion in the nuclear pore complex of live cells. Proc. Natl. Acad. Sci. U. S. A. 109 (25), 9863-9868 (2012).
- Sanchez, S. A., Tricerri, M. A., Gratton, E. Laurdan generalized polarization fluctuations measures membrane packing micro-heterogeneity in vivo. Proc. Natl. Acad. Sci. U. S. A. 109 (19), 7314-7319 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Fluorescence correlation spectroscopy of intact nuclear pore complexes. Biophys. J. 101 (4), 27-29 (2012).
- Di Rienzo, C., et al. Unveiling LOX-1 receptor interplay with nanotopography: mechanotransduction and atherosclerosis onset. Sci. Rep. 3, 10-1038 (2013).
- Unruh, J. R., Gratton, E. Analysis of molecular concentration and brightness from fluorescence fluctuation data with an electron multiplied CCD camera. Biophys. J. 95 (11), 5385-5398 (2008).
- Kannan, B., et al. Electron multiplying charge-coupled device camera based fluorescence correlation spectroscopy. Anal. Chem. 78 (10), 3444-3451 (2006).
- Jones, S. A., Shim, S. H., He, J., Fast Zhuang, X. three-dimensional super-resolution imaging of live cells. Nat. Methods. 8 (6), 499-508 (2011).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy. 3 (10), 793-795 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Hess, S. T., Girirajan, T. P., Mason, M. D. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys. J. 91 (11), 4258-4272 (2006).
- Manley, S., et al. High-density mapping of single-molecule trajectories with photoactivated localization microscopy. Nat. Methods. 5 (2), 155-157 (2008).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Klar, T. A., Hell, S. W. Subdiffraction resolution in far-field fluorescence microscopy. Opt. Lett. 24 (14), 954-956 (1999).
- Eggeling, C., et al. Direct observation of the nanoscale dynamics of membrane lipids in a living cell. Nature. 457 (7233), 1159-1162 (2009).
- Hedde, P. N., et al. Stimulated emission depletion-based raster image correlation spectroscopy reveals biomolecular dynamics in live cells. Nat. Commun. 4, Forthcoming.
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophys. J. 88 (5), 3601-3614 (2005).
- Brown, C. M., et al. Probing the integrin-actin linkage using high-resolution protein velocity mapping. J. Cell Sci. 119, 5204-5214 (2006).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proc. Natl. Acad. Sci. U. S. A. 110 (30), 12307-12312 (2013).
- Mueller, V., et al. STED nanoscopy reveals molecular details of cholesterol- and cytoskeleton-modulated lipid interactions in living cells. Biophys. J. 101 (7), 1651-1660 (2011).
- Kleusch, C., Hersch, N., Hoffmann, B., Merkel, R., Csiszar, A. Fluorescent lipids: functional parts of fusogenic liposomes and tools for cell membrane labeling and visualization. Molecules. 17 (1), 1055-1073 (2012).
- Ries, J., Chiantia, S., Schwille, P. Accurate determination of membrane dynamics with line-scan FCS. Biophys. J. 96 (5), 1999-2008 (2009).
- Kolin, D. L., Wiseman, P. W. Advances in image correlation spectroscopy: measuring number densities, aggregation states, and dynamics of fluorescently labeled macromolecules in cells. Cell Biochem. Biophys. 49 (3), 141-164 (2007).
- Digman, M. A., et al. Measuring fast dynamics in solutions and cells with a laser scanning microscope. Biophys. J. 89 (2), 1317-1327 (2005).
- Ritchie, K., et al. Detection of non-Brownian diffusion in the cell membrane in single molecule tracking. Biophys. J. 88 (3), 2266-2277 (2005).
- Voie, A. H., Burns, D. H., Spelman, F. A. Orthogonal-plane fluorescence optical sectioning: three-dimensional imaging of macroscopic biological specimens. J. Microsc. 170, 229-236 (1993).
- Huisken, J., Swoger, J., Del Bene,, Wittbrodt, F., J,, Stelzer, E. H. Optical sectioning deep inside live embryos by selective plane illumination microscopy. Science. 305 (5686), 1007-1009 (2004).
- Wohland, T., Shi, X., Sankaran, J., Stelzer, E. H. Single plane illumination fluorescence correlation spectroscopy (SPIM-FCS) probes inhomogeneous three-dimensional environments. Opt. Express. 18 (10), 10627-10641 (2010).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}