

Abstract
Het is steeds duidelijker dat de ruimtelijke verdeling en de beweging van membraan componenten zoals lipiden en proteïnen zijn de belangrijkste factoren in de regulatie van vele cellulaire functies geworden. Vanwege de snelle dynamiek en kleine structuren betrokken zijn, een zeer hoge ruimtelijke resolutie moet het werkelijke gedrag van moleculen te vangen. Hier presenteren we de experimentele protocol voor het bestuderen van de dynamica van fluorescent gelabelde plasma-membraaneiwitten en lipiden in levende cellen met hoge spatiotemporele resolutie. Met name is deze benadering niet te elk molecuul volgen, maar berekent gedrag populatie met alle moleculen in een bepaalde regio van het membraan. Uitgangspunt is een snelle beeldvorming van een bepaald gebied op het membraan. Daarna wordt een volledige spatio-temporeel autocorrelatiefunctie berekend correlatie verkregen beelden met toenemende vertraging, bijvoorbeeld elke 2, 3, n herhalingen. Het kan aangetoond worden dat de breedtevan de piek van de ruimtelijke autocorrelatiefunctie toeneemt met toenemende vertraging als functie van deeltjesbeweging door diffusie. Derhalve passen de reeks autocorrelatiefuncties maakt uittreksel het werkelijke eiwit mean square verplaatsing van imaging (iMSD), hier voorgesteld in de vorm van schijnbare diffusiecoëfficiënt versus gemiddelde verplaatsing. Dit levert een kwantitatief beeld van de gemiddelde dynamica van enkele moleculen met nanometer nauwkeurigheid. Door een GFP-gelabelde variant van de transferrinereceptor (TfR) en een ATTO488 gemerkt 1-palmitoyl-2-hydroxy-sn-glycero-3-fosfoethanolamine (PPE) is het mogelijk om de spatio-temporale regulering van eiwitten en lipiden diffusie observeren urn sized membraan gebieden in de micro-naar milli-seconde tijdbereik.
Introduction
Uitgaande van de originele "vloeistof mozaïek"-model door Singer en Nicolson, heeft het beeld van cellulaire plasmamembraan voortdurend bijgewerkt tijdens de laatste decennia om de opkomende rol van het cytoskelet en lipidedomeinen 1,2 omvatten.
De eerste waarnemingen werden verkregen door TL herstel na fotobleken (FRAP) onthulling dat een aanzienlijk deel van membraaneiwitten immobiel is 3-5. Deze baanbrekende studies, hoewel zeer informatief, last van de relatief slechte resolutie in ruimte (micron) en de tijd (in seconden) van FRAP setups. Ook wordt een ensemble gemiddelde meting, FRAP ontbreekt in het geven van informatie over enkele molecule gedrag.
In deze context, de mogelijkheid specifiek label een molecuul met zeer lichte markeringen (waardoor de studie van het diffusieproces een molecuul tegelijk) is zeer succesvol geweest. In het bijzonder, door op detijdsresolutie van de Single Particle Tracking (SPT) benadering van de microseconde tijdschaal, Kusumi, ea. toegang gekregen tot onbekende eigenschappen van lipiden en eiwitten dynamiek die in belangrijke mate bijgedragen aan de erkenning van de rol van actine gebaseerde membraan skelet in membraan fysiologie 6 , 7. Deze bevindingen gegenereerd zogenaamde het "paal en hek-model, waarbij lipiden en eiwitten diffusie wordt geregeld door actine gebaseerde skelet. Echter, om de toegang tot de enorme hoeveelheid door NBP vele experimentele Bull informatie moet worden aangepakt. In het bijzonder wordt de labelingsprocedure normaliter samengesteld door vele stappen, zoals productie, zuivering en invoering van de gemerkte species in het systeem. Bovendien grote labels, zoals quantum dots of metalen nanodeeltjes, zijn vaak nodig om de sub-milliseconde tijdschaal en de verknoping van de doelgroep moleculen door het label kon niet vermeden worden in veel gevallen te bereiken. Tenslotte vele trajectenmoeten worden opgenomen statistische criteria passen en gelijktijdig een lage dichtheid van het label moet volgen mogelijk.
Vergeleken met SPT, fluorescentie correlatie spectroscopie (FCS), overwinnen veel van deze nadelen is een veelbelovende benadering voor moleculaire dynamica te bestuderen. In feite, FCS werkt goed ook zwak en dichte labels, waardoor de dynamiek van fluorescerend proteïne gemerkte moleculen te bestuderen in transient getransfecteerde cellen. Ook kun bereiken hoog statistieken een beperkte tijd. Tot slot, ondanks de "hoge" dichtheid van labels FCS biedt enkele moleculen informatie. Door al deze eigenschappen, FCS staat voor een zeer rechtlijnige aanpak en is uitgebreid toegepast op lipiden en eiwitten dynamiek te bestuderen, zowel in model membranen en in levende cellen 8-10. Vele verschillende benaderingen zijn voorgesteld om het vermogen van FCS om de details van moleculaire diffusie onthullen verhogen. Zo was sheigen, dat door het uitvoeren van FCS op verschillende afmetingen observatie gebieden kan men een "FCS diffusie wet" verhelderend verborgen eigenschappen van moleculaire beweging 11,12 definiëren. Naast het feit dat varieerde in grootte, werd het aandachtsgebied ook gedupliceerd 13, verhuisde in de ruimte langs de lijnen 14-20 of geconjugeerd met snelle camera's 21,22. Met behulp van deze 'ruimte-tijd' correlatie benaderingen, werden relevante biologische parameters van verschillende membraan componenten kwantitatief beschreven op zowel modelmembranen en werkelijke biologische degenen, waardoor aldus inzicht in de membraan-ruimtelijke organisatie.
In alle FRAP en FCS toepassingen dusver beschreven de grootte van het concentratiegebied is een limiet ruimtelijke resolutie die niet kan worden overwonnen. Verschillende beeldvormende technieken super-resolutie zijn recent ontwikkeld om deze limiet te omzeilen. Sommige zijn gebaseerd op lokalisatie precisie, zoals stochastische optische reconstructie microscopie (STORM) <sup> 23,24, fotoactivering lokalisatie microscopie (PALM) 25, fluorescentie PALM (FPALM) 26 en enkele deeltjes volgen PALM (sptPALM) 27: de relatief grote hoeveelheid fotonen vereist op elke foto, beperkt echter de tijdsresolutie van deze methoden om ten minste enkele milliseconden, waardoor hun toepasbaarheid in vivo belemmeren.
Daar staat tegenover dat een veelbelovend alternatief voor de super-resolutie imaging geopend door ruimtelijk moduleren van de fluorescentie-emissie met gestimuleerde emissie uitputting methoden (STED of omkeerbare verzadigbaar optische fluorescentie overgangen (RESOLFT)) 28,29. Deze benaderingen combineren de vormgeving van de waarneming volume ruim onder de diffractie limiet de mogelijkheid om snelle scanning microscopen en detectiesystemen gebruikt. In combinatie met fluorescentie fluctuatie analyse, STED microscopie toegestaan om direct sonde nanoschaal spatiotemporele dynamiek van lipiden en proteins in levende celmembranen 30,31.
Dezelfde fysische hoeveelheden STED gebaseerde microscopie kan worden verkregen door een gemodificeerde spatio-temporeel beeld correlatie spectroscopie (PEN 32,33) methode die geschikt is voor de studie van de dynamiek van fluorescent-gelabelde membraaneiwitten en / of lipiden in levende cellen en door een commerciële microscoop. Het experimentele protocol hier gepresenteerd is samengesteld door enkele stappen. De eerste vereist een snelle beeldvorming van het gebied van belang op het membraan. Vervolgens wordt de resulterende stapel beelden worden gebruikt om de gemiddelde spatiotemporeel correlatiefuncties berekenen. Door toepassing van de reeksen van correlatiefuncties de moleculaire diffusie recht "kan direct vanuit beeldvorming verkregen in de vorm van een schijnbare diffusiecoëfficiënt (D app) - vs -gemiddelde verplaatsingsplot. Dit perceel kritisch afhankelijk van de omgeving verkend door de moleculen en maakt direct de werkelijke verspreiding modi herkennenvan het lipide / eiwit plaats.
In meer details, zoals eerder aangegeven 34, de tijd-ruimtelijke auto-correlatie-functie van de verworven afbeelding serie kritisch afhankelijk van de dynamiek van de moleculen bewegen in het verzamelde beeld-serie (let op dat dezelfde redenering in een lijn overname kan worden toegepast waar slechts een dimensie in de ruimte wordt beschouwd). In het bijzonder definiëren we de correlatiefunctie als:
 (1)
(1)
waar  vertegenwoordigt de gemeten fluorescentie-intensiteit in de positie x, y en op tijdstip t,
vertegenwoordigt de gemeten fluorescentie-intensiteit in de positie x, y en op tijdstip t, ![]() en
en ![]() is de afstand in de x enrespectievelijk de y-richting,
is de afstand in de x enrespectievelijk de y-richting, ![]() vertegenwoordigt het tijdsverloop, en
vertegenwoordigt het tijdsverloop, en ![]() vertegenwoordigt het gemiddelde. Deze functie kan worden uitgedrukt als:
vertegenwoordigt het gemiddelde. Deze functie kan worden uitgedrukt als:
 (2)
(2)
waarbij 'N' staat voor het gemiddelde aantal moleculen in de observatie gebied, ![]() is de convolutie-bewerking in de ruimte, en
is de convolutie-bewerking in de ruimte, en  vertegenwoordigt de autocorrelatie van de instrumentale taille. Dit laatste kan worden opgevat als een maat voor hoe de fotonen van een emitter zijn verspreid in de ruimte door de optische / opname opstart (de zogenaamde puntspreidingsfunctie, PSF, generally goed benaderd door een Gauss-functie). Tenslotte
vertegenwoordigt de autocorrelatie van de instrumentale taille. Dit laatste kan worden opgevat als een maat voor hoe de fotonen van een emitter zijn verspreid in de ruimte door de optische / opname opstart (de zogenaamde puntspreidingsfunctie, PSF, generally goed benaderd door een Gauss-functie). Tenslotte  is de kans om een deeltje te vinden op afstand
is de kans om een deeltje te vinden op afstand ![]() en
en ![]() na een bepaalde tijd
na een bepaalde tijd ![]() . Als we kijken naar een diffusief dynamiek, waarin deeltjes bewegen willekeurig in alle richtingen en de netto fluxen niet aanwezig zijn, is deze functie ook goed benaderd door een Gauss-functie waarbij de variantie kan worden geïdentificeerd als de Mean Square Verplaatsing (MSD) van het bewegende deeltje . Aldus, de taille van de correlatiefunctie (ook aangeduid als
. Als we kijken naar een diffusief dynamiek, waarin deeltjes bewegen willekeurig in alle richtingen en de netto fluxen niet aanwezig zijn, is deze functie ook goed benaderd door een Gauss-functie waarbij de variantie kan worden geïdentificeerd als de Mean Square Verplaatsing (MSD) van het bewegende deeltje . Aldus, de taille van de correlatiefunctie (ook aangeduid als ![]() ), Kan worden gedefinieerd als de som van het deeltje MSA en instrumentele taille en kan worden gemeten door een Gauss fitting van de correlatie functie voor elke vertraging. De gemeten i MSD kan worden gebruikt om een schijnbare diffusiecoëfficiënt van de bewegende moleculen berekenen
), Kan worden gedefinieerd als de som van het deeltje MSA en instrumentele taille en kan worden gemeten door een Gauss fitting van de correlatie functie voor elke vertraging. De gemeten i MSD kan worden gebruikt om een schijnbare diffusiecoëfficiënt van de bewegende moleculen berekenen ![]() en een gemiddelde verplaatsing
en een gemiddelde verplaatsing ![]() zoals:
zoals:
 (3)
(3)
 (4)
(4)
Enkele overwegingen op experimentele opstelling gebruikt kan de lezer begeleiden gedurende de volgende secties. Om de fluoroforen selectief exciteren van het basale membraan van levende cellen zullen we een totale interne reflectie (TIR) belichting, onder gebruikmaking van een commercieel TIR fluorescentie (TIRF) microscoop (vindt u bij materialen). Bovendien, teneinde th verzamelene fluorescentie zullen we een sterke vergroting doelstelling gebruiken (100X NA 1,47, wordt hoge numerieke apertuur nodig TIRF verlichting) en een EMCCD camera (fysieke grootte van de pixel op de chip 16 urn). Een pixelgrootte van 100 nm te bereiken passen we een extra vergroting lens van 1,6 x. Zoals hieronder besproken, zou een tijdresolutie dan 1 msec nodig om de dynamiek van snelle membraanlipiden goed beschreven onder 100 nm. Om deze tijdresolutie bereiken moeten we een van belang (ROI) te selecteren kleiner dan de gehele chip van de camera (512 x 512). Op deze manier zal de camera een beperkt aantal regels verhogen de tijdsresolutie lezen. In dit regime uitlezing geschikte lijst beperkte duur door de benodigde tijd en is meestal in de orde van milliseconden 512 x 512 pixel EMCCD tot kosten van de blootstelling verschuiven naar de uitlezing chip op de camera. Om deze limiet te verslaan, een opkomende technologie maakt het verschuiven van de ROI-lijn in plaats van het gehele frame, wet een praktische effectieve vermindering van de blootgestelde chip grootte (genaamd Cropped Sensor Mode in onze EMCCD). Voor deze configuratie om effectief te zijn, moet de chip buiten de ROI worden gedekt door een paar spleten gemonteerd in het optische pad. Dankzij deze opstelling een tijdresolutie tot 10 -4 seconden worden bereikt. Let er echter op dat deze aanpak kan worden gekoppeld met veel verschillende experimentele opstellingen, zoals beschreven in de paragraaf 'discussie'.
Demonstratie van de werkwijze wordt voorzien in levende cellen met zowel een ATTO488 gemerkt 1-palmitoyl-2-hydroxy-sn-glycero-3-fosfoethanolamine (ATTO488-PPE) en een GFP-gelabelde variant van de transferrinereceptor (GFP TfR). Bij ATTO488-PPE kan deze benadering succesvol een nagenoeg constant D app als functie van gemiddelde verplaatsing aangeeft een meestal vrij diffusie herstellen, zoals eerder gemeld 30,35. Daarentegen TfR-GFP vertoont een afnemende D
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1 systeeminmeting
- Point Spread Function (PSF) kalibratie
- Verdun 10 pi van 30 nm fluorescente bead oplossing (ongeveer 5 uM) in 90 pl gedestilleerd water en vervolgens ultrasone trillingen de oplossing gedurende 20 minuten. Snij een vierkant (1 cm x 1 cm) stuk agarose gel (3%) en borg 10 ul van de oplossing op de gel. Overturn het stukje gel op de bodem glas van een 2 cm petrischaal en knijp de druppel op het glas.
- Zet de overname setup, zet het monster in de houder, zet de juiste belichting en EMgain (100 msec en 1000 zijn goed, maar de parameters te optimaliseren volgens het systeem) en wacht tot de camera is afgekoeld.
- Stel de camera blootstelling aan 100 msec, camera EMgain 1000, overname-modus om Transfer, 100 herhaling en auto Lijst opslaan instellen.
- Met behulp van het oculair en doorgelaten licht focus op de grens van de gel en vervolgens de doelstelling om het centrum van de gel, past de focus en start de laser uitlijning procedure (in LAS AF, selecteer 'TIRF setup' en volg de automatische uitlijning procedure).
- Zoek een gezichtsveld met enkele, geïsoleerde plekken, nauwkeurig richten op de heldere plek (die meestal vertegenwoordigt kralen aggregaat) als referentie, de overname van 100 frames en herhaal de stappen 5-6 keer om verschillende single plekken te verwerven.
- Importeer de verworven-serie met een gegevensverwerkende programma en het gemiddelde van de stapel in de tijd (figuur 1A) en selecteer een enkele geïsoleerde kraal. Zorg ervoor dat de kleinste deeltjes te selecteren om aggregaten te voorkomen.
- Passen bij de gekozen intensiteit distributie (een voorbeeld van enkele kralen profiel wordt weergegeven in Figuur 1B) met een Gauss-functie met behulp van het commando "gaussfit" (in de ICS-Matlab gereedschappen in de Materials in Matlab). Controleer de goedheid van de pasvorm door de inspectie van de verkregen residuen (een voorbeeld van een ingerichte Gauss-profiel met de bijbehorende resten wordt gepresenteerd in Figuur 1B).
- Camerakalibratie
- Schakel de camera in en wacht tot de camera is afgekoeld. Stel de camera overname instelling, (dat wil zeggen, voor de gebruikte camera zetten we de blootstelling aan 0,5 msec, camera EMgain 1000, overname-modus om Cropped Mode, de ROI grootte tot 32 x 128, 10.000 herhalingen) en start de overname van de achtergrond camera signaal.
- Import verworven frame reeks met een gegevensverwerkende programma. Bereken en inspecteer de gemiddelde intensiteit van elke pixel in om te controleren of de achtergrond camera is ongeveer plat in het geselecteerde gebied van de chip. In bijgesneden modus verwijdert de eerste en de laatste horizontale lijnen (3 tot 10 afhankelijk van de grootte van de ROI) voor elk frame, omdat de achtergrond camera gewoonlijk voorgespannen in de grenslijnen.
- Maak een histogram van de waarden (ook gedefinieerd Digital Level, DL) in de verkregen beelden stack (met de opdracht 'gesch "in Matlab) en plot de logaritmevan de resulterende frequentie (met semilogy commando in Matlab). Een voorbeeld van DL distributiesysteem voor camera achtergrond wordt weergegeven in figuur 2.
OPMERKING: Als de camera goed werkt, zal het perceel een ongeveer Gaussische piek (een parabolische profiel in log-schaal) die de verdeling van de waarden die op nul foton gevolgd door een exponentiële verval (een lijn met negatieve helling in log-schaal tonen ) de verdeling van de waarden die 1 foton (figuur 2) staat. Vooral het midden en de variantie van de Gaussische functie vertegenwoordigen de camera offset and error respectievelijk terwijl de vervalconstante van het exponentiële deel vertegenwoordigt een schatting van de DL toegewezen door de camera elke foton. In Matlab gebruiken de sectie "CalibrateCamera" van het script in ondersteunende materialen. - Herhaal deze handeling voor alle geselecteerde Camera EMGain en Gain.
2 gelabeldeCell Bereiding
- Om de voor lipide bijmenging 36 liposomen Hiertoe wordt afzonderlijk 1 mg DOPE (1,2-dioleoyl- sn glycero-3-fosfoethanolamine), 1 mg van DOTAP (1,2-dioleoyl-3-trimethylammonium-propaan), en 1 mg van de PPE-ATTO488 in 1 ml chloroform. Meng 0,5 ml DOPE-oplossing, 0,5 ml DOTAP oplossing en 25 gl PPE-ATTO488 oplossing en droog onder vacuüm gedurende 24 uur. Voeg 0,5 ml HEPES-buffer 20 mM, vortex gedurende 15 min en ultrasone trillingen gedurende 15 minuten bij 40 ° C.
- Om de cel te bereiden, was 3 keer met PBS een p100 gerecht van samenvloeiende CHO-K1 (Chinese hamster ovarium), voeg 1 ml trypsine en bewaar in de incubator gedurende 5 minuten. Suspendeer vrijstaande cellen toevoegen 9 ml DMEM / F12 medium aangevuld met 10% FBS en zaden 150 gl celoplossing in een petrischaal met 800 ul van hetzelfde medium.
- Store in incubator gedurende 24 uur bij 37 ° C en 5% CO2. Voor lipide incorporatie, vervang cel medium met 500gl serumvrij medium; Na 30 min, voeg 2 ui liposomen oplossing; na 15 min wassen met PSB en voeg nieuwe DMEM / F12 medium voor beeldvorming.
- Voor transfectie, transfecteren cellen volgens Lipofectamine protocol (instructies fabrikant) met TfR-GFP plasmide en winkel 24 uur in de incubator voor de beeldvorming.
3 Data Acquisition
- Voorbereiding setup
- Om de microscoop thermostaat, 24 uur voor het experiment zet de incubator.
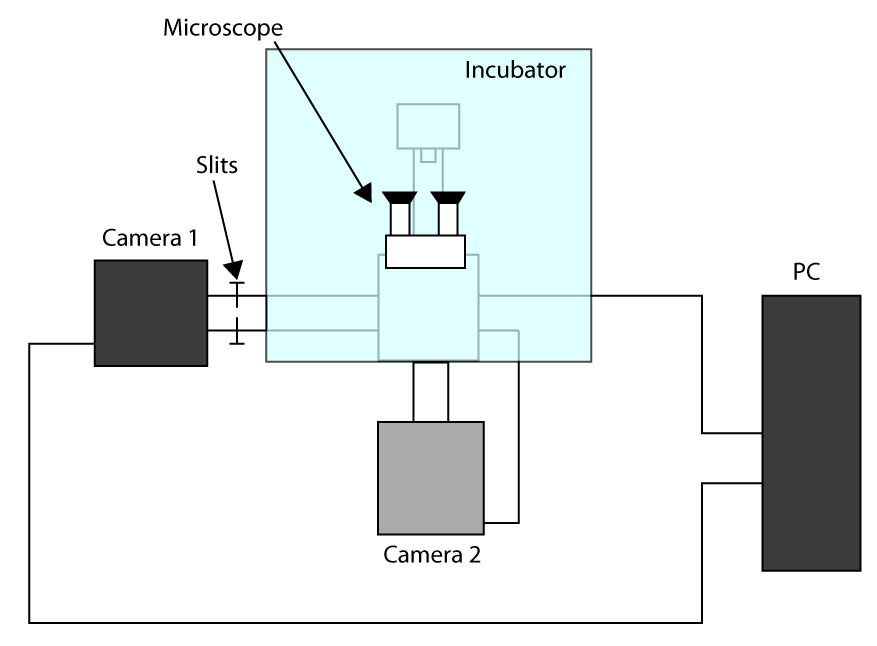
- Om de snelste haalbare acquisitietijd toepassing werken Vrijstaande Sensor Mode (zie inleiding) en gebruik een eerste camera voor afbeelden (camera 1) en tweede camera voor de cel (2 camera) selecteren. Een schema van de installatie configuratie wordt gepresenteerd in aanvullende Figuur S1. Om vervolgens de twee camera inschakelen van de microscoop richten en wacht tot de camera afkoelen.
- Stel op beide camera's de parameters voor het doorgelaten licht beeldvorming (dwz
- Zet de monsters in de houder en scherpstellen met oculair, stuurt het licht naar de camera 1 en duw de spleten waardoor het licht alleen op de ROI voor cell imaging (hier een 32 x 32 pixels ROI).
- Verplaats een cel in het geselecteerde gebied en sturen het licht naar de camera 2, dan trek je een ROI in de software die camera 2 beheersen om een verwijzing te hebben.
- Imaging (figuur 3A)
- Allereerst richt de TIRF laser volgens de procedure van je setup. In onze setup, selecteer de 'TIRF setup' en start de automatische uitlijning procedure. Wanneer de laser is uitgelijnd ingesteld 70 nm penetratiediepte (ongeveer 70 °).
- Stel belichtingstijd tot 70 msec en EMGain tot 100 op beide camera 1 en camera 2; Vervolgens selecteert u een cel met behulp van camera 1, stuur dan het licht op camera 2 en nauwkeurig richten van de celmembraan. Stel de minimale blootstelling op camera 2, 1000 EMGain, Cropped Sensor Mode, 10 5 herhalingen en stel autosave zo past bestanden (Flexible Image Transport System, een formaat dat gemakkelijk kan worden beheerd).
- Start de overname om het beeld-serie op te nemen. Laat de Gain en de bijgesneden Mode op temperatuur kan stabiliseren voordat u tot aanschaf van een nieuwe cel, herhaal de laatste twee stappen om 8-10 cellen verwerven.
4 Berekening van de Mean Square Verplaatsing van Imaging (i MSD)
Opmerking: Het volgende protocol kan direct worden toegepast op ruwe data. Tegelijkertijd, het hele protocol geldt voor data acquisities gesimuleerd zowel in Matlab en in SimFCS. De link naar de bijbehorende tutorials te vinden in de sectie 'Materialen'.
- Berekening van Matlab
- Importeer de verworven serie Matlab met behulp ImportImageSeries script. Bereken de gemiddelde intensiteit van elk beeld in de tijd met behulp van de command betekent op de eerste 2 afmetingen en gebruik plot de resulterende vector te bekijken.
- Indien meer dan 10% van fotobleken aanwezig is, gooi de reeks of verwijder het eerste deel daarvan. Als het lager is, proberen om het effect op de correlatie functie te corrigeren door het aftrekken van elke afbeelding zijn gemiddelde intensiteit, zoals vóór 37.
- Bereken de gemiddelde intensiteit van elke pixel met de gemiddelde op de derde dimensie zien resulterende beeld.
OPMERKING: Bijzondere aandacht is vereist om artefacten correlaties voorkomen. In feite, zoals eerder aangegeven voor soortgelijke technieken 38, Celranden evenals onscherp blaasjes kan een sterke correlatie te introduceren. Indien de inspectie van de gemiddelde afbeelding onthult cel grenzen of onscherp blaasjes, proberen de betrokken regio de overname anders gooi te sluiten. Om het effect van deze onbeweeglijke structuren corrigeren aftrekken van de gemiddelde tijdelijke intensiteit van elke pixel 39. - Bereken thij tijdruimtelijk correlatie (G (ξ, χ, τ)) met behulp van de functie CalculateSTICScorrfunc. Verwijder G (ξ, χ, 0), omdat de correlatie te wijten aan de opname ruis bij weinig licht regime domineert G (0,0,0); de correlatie te wijten aan de detector domineert de G (± 1,0,0), en deeltjes bewegen tijdens de belichtingstijd G (ξ, χ, τ) voor τ = 0 kan vervormen door het verhogen van de gemeten taille (dit effect verdwijnt voor τ > 0) 34.
- Gemiddeld G (ξ, χ, τ> 0) met behulp van een logaritmische tijd-bin om het lawaai te verminderen door het gebruik van de functie "LogBinStack" in Steunmateriaal en dan passen de resulterende G (ξ, χ, τ) met behulp van de functie "gaussfit" van de ICS-Matlab gereedschappen in de Materials op de i MSD (de tweede kolom van de resulterende array) te herstellen.
- Zet de verkregen taille σ (τ) 2 (i MSD) als functie van de tijd. Als de gegevens zijn te luidruchtig, probeer dan om het aantal acqu verhogenired frames, verhogen het laservermogen, gemiddeld meer G (ξ, χ, τ) samen.
- Berekening door SimFCS
- Open de verworven-bestanden met behulp van ImageJ BioFormat importeur plugin en sla verworven series als Tiff volgorde.
- Open SimFCS en selecteer RICS gereedschap en kies Bestand> Import Meerdere beelden (aanvullende figuur S2).
- Selecteer Fit, plaatst u de juiste acquisitie parameters en sluit de pasvorm venster (aanvullende figuur S3).
- Selecteer Weergave> Gemiddeld Intensiteit> CH1 en verifiëren van de aanwezigheid van fotobleken (aanvullende figuur S4).
- Indien meer dan 10% van fotobleken aanwezig discard de reeks of het mogelijk verbruiker het beeld sequentie verwijderen van het eerste deel van de reeks.
- Als bleken het lager is dan 10% selecteert u Extra> i MSD> Set Parameters, check 'Gebruik voortschrijdend gemiddelde', gelegen in de ROI-paneel aan de linkerzijde eenantal frame voor het voortschrijdend gemiddelde Let op de correspondent moment hoger is dan de karakteristieke diffusie tijd (voor deeltjes bewegen met 1 urn 2 sec -1 een tijd van 10 seconden is een bewegend gemiddelde)
- Selecteer Extra> iMSD> Bereken i MSD (aanvullende figuur S5) en fit en exporteren de i MSD uit de memo pad (aanvullende figuur S6).
5 Berekening van de Diffusion wet van de i MSD
- Plaats de eerste paar punten aan het intercept (σ 0 2) extrapoleren (5 punten zijn meestal genoeg, maar meer punten kunnen worden geplaatst indien een lineair gedrag vertonen) en vergelijk deze waarde met de eerder gemeten PSV 2. Als ze vergelijkbaar zijn, zijn de dynamiek van geïsoleerde fluoroforen gevolgd. Indien daarentegen σ 0 2 >> PSV 2 proberen te verwerven sneller om ervoor te zorgen datgeen verborgen dynamiek aanwezig 34.
- Bereken de schijnbare diffusiecoëfficiënt (D app) en de gemiddelde verplaatsing (R) met de vergelijkingen 3 en 4 (zie inleiding).
- Plot D app als functie van R een diffusie wet vergelijkbaar met wat gemeten spot variatie gebaseerd FCS 12 (figuur 3D) te verkrijgen.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Om instrumentele taille kalibreren kan het beeld van een fluorescent nano-bead maatregel zoals beschreven in stap 1.1 Protocol. Een typische fluorescerende beeld van deze kralen is weergegeven in figuur 1. Fitting volgens intensiteitsverdeling van een 2D Gaussische functie teruggeeft goede residuen en laat meten instrumentale taille bij 270 nm. Deze waarde komt goed overeen met de verwachte diffractielimiet geschat door de Rayleigh vergelijking. Deze kalibratie is niet nodig voor de meting van het dynamische, maar het vereist is de schijnbare deeltjesgrootte te meten.
Een typische frequentieverdeling van camera achtergrond is weergegeven in figuur 2. De piek bij ongeveer 180 DL is door de camera reactie geen foton, en vertegenwoordigt de bijdrage van analoge Digital (AD) converter. Deze bijdrage kan worden benaderd als een Gauss-verdeling om een schatting van de offset en de variantieingevoerd bij de signaalopname. Boven 200 DL het digitale niveau distributie wordt exponentiële (lineair in logaritmische schaal) en vertegenwoordigt de gemiddelde camera reactie op een enkel foton. Koppeling dit deel met een exponentiële verdeling maakt de metingen van de gemiddelde DL toegewezen aan elke foton. Hoe hoger de verhouding tussen de gemiddelde DL toegewezen aan elke foton en de AD converter fout, hoe lager de ruis in de berekende correlatiefunctie zijn. Bovendien is de gemiddelde foton reactie maakt de schatting van de camera dynamisch bereik.
Een diagram van de volledige experimentele procedure wordt samengevat in figuur 3 en een afbeelding van Atto488 PPE-insertie in het membraan is weergegeven in figuur 4A. Een vertegenwoordiger TIRF beeld van de basale membraan van CHO-cellen gelabeld met Atto488-PPE is weergegeven in figuur 4B. Verschillende heldere vlekken aanwezig buiten de cel door LIposomes gestapeld op het glas. Ze kunnen worden verwijderd op een ROI geselecteerd op wandgedeelte overwegend uniform in fluorescentie (dwz. De cellulaire plasmamembraan). Zoals verwacht de gemeten diffusie wet (figuur 4C) van dit lipide is vlak, wat wijst op een meestal gratis verspreiding, zoals eerder aangegeven door STED-FCS metingen 30,35. Het is vermeldenswaard dat alle getoonde verplaatsing waarden zijn lager dan de diffractie limiet, met duidelijke vermelding van het vermogen van deze benadering van de moeite super-oplossing gemiddelde moleculaire verplaatsingen ruim onder de diffractie limiet en omlaag om enkele tientallen nanometers.
Een schematisatie van TfR-GFP dimeer insertie in het membraan is weergegeven in figuur 5A. Vele studies toonden dat de cytoplasmatische staart van de receptor interageert met het membraan skelet, dat op zijn beurt fungeert als een hek voor de receptor mobiliteit 12,40. Een vertegenwoordiger TIRF beeld van een CHO-cel die TfR-GFP is presented in figuur 5B. Lage fluorescentie-intensiteit cellen de voorkeur, als het membraan dichter bij de oorspronkelijke toestand en de waarschijnlijkheid van voorwerpen in verband met de overexpressie wordt geminimaliseerd. Bovendien moet het centrale deel van de cel worden vermeden, omdat de effecten van out-of-focus fluorescentie (van cytoplasma, bijvoorbeeld) aanwezig zijn. Zoals verwacht gemeten diffusie wet (figuur 5C) voor TfR-GFP een eerste vlak gedrag onder 100 nm, met een gemiddelde D app van ongeveer 0,7 urn 2 sec -1, gevolgd door daaropvolgende snelle daling duidelijk diffusie tot 0,2 urn 2 sec -1 (de waarde meestal gemeten door buigingsbegrensde FCS 12). Dit resultaat toont aan dat onze benadering gemakkelijk meten de gemiddelde verplaatsing van GFP gemerkte eiwitten met een resolutie van enkele tientallen nanometers. Bovendien is de ruimtelijke schaal waarop D app begint te sets de karakteristieke verlagenruimtelijke schaal van eiwit gedeeltelijke opsluiting door het membraan skelet rond 120 nm, in overeenstemming met eerdere schattingen 6.

Figuur 1 Kalibratie van Point Spread Function. (A) pseudokleur beeld van een geïsoleerde kralen en parels aggregaten. (B) 3D grafiek van de intensiteit profiel van een geïsoleerd kraal toont een welbepaald profiel Gaussiaans. (C) Fit van de intensiteitsverdeling van een Gaussische functie (bovenste paneel) met de overeenkomstige residuen (onderste paneel). De goede overeenkomst tussen de gemonteerde distributie en de gemeten intensiteit profiel is ook een bewijs dat de instrumentale PSF kan worden benaderd door een Gaussische functie. Klik hier om vi ew een grotere versie van deze figuur.

Figuur 2 Kalibratie van Camera reactie op enkele fotonen. De figuur toont de Digital Level (DL) distributie voor camera achtergrond in een 32 x 128 ROI, blootstelling 0,5 msec, in Cropped Sensor Mode. De piek bij ongeveer 180 dl de camera reactie geen fotonen. In het bijzonder vertegenwoordigt de bijdrage van de analoge Digital (AD) converter en kan worden benaderd met een Gaussische functie te schatten de offset en de variantie die door het signaalopname. Boven 200 DL de distributie van digitale niveaus wordt exponentieel, vertegenwoordigt de gemiddelde camera reactie op een enkel foton. Het meten van deze parameters maakt het schatten van de dichtheid van fotonen die zijn opgenomen tijdens de overname.s / ftp_upload / 51994 / 51994fig2highres.jpg "target =" _blank "> Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 3 Schematisering van de werkwijze. (A) Breed beeldvormend EMCCD camera wordt toegepast onder-milliseconde bereikt, terwijl TIRF microkopie wordt benut om nauwkeurige optische sectie van het plasmamembraan verschaffen. (B) De resulterende stapel afbeeldingen wordt autocorrelated om de gemiddelde ruimtelijke berekenen -temporal correlatie functie. Deze correlatie functie goed benaderd door een Gaussische functie (zie inleiding) en verspreidt op tijd op basis deeltje verplaatsingen. (C) Om derhalve de kwantificeren verspreiding van de correlatiefunctie door moleculaire verdringing, passend met een Gauss ian functie wordt uitgevoerd. Dit maakt meting van de moleculaire diffusie law "rechtstreeks van beeldvorming, in de vorm van schijnbare diffusiecoëfficiënt versus gemiddelde verplaatsingsplot. (D) Met deze curve kan moleculaire diffusie functies direct worden geïdentificeerd zonder de noodzaak van een interpretatieve model of aannamen over de ruimtelijke ordening van het membraan. In feite, kan vrij diffunderen moleculen een constante schijnbare diffusiviteit weergegeven als hun mobiliteit niet afhankelijk van de ruimtelijke omvang van de meting. Daarentegen zal gedeeltelijke afsluiting moleculen een vrij constante schijnbare diffusie weer voor verplaatsingen kleiner dan opsluiting grootte, dan is een afnemende diffusiviteit voor ruimtelijke schalen groter zijn dan opsluiting grootte. Aldus kan het uiterlijk van een verlaging van de schijnbare diffusiecoëfficiënt worden uitgelegd als een vingerafdruk van voorbijgaande opsluiting, terwijl de bijbehorende ruimtelijke schaal kan worden gebruikt om de ruimtelijke uitbreiding van de bevalling. .jove.com / files / ftp_upload / 51994 / 51994fig3highres.jpg "target =" _blank "> Klik hier om een grotere versie van deze afbeelding te bekijken.

Figuur 4 ATTO488 PPE-diffusie TIRF beeld van CHO basaalmembraan gelabeld met ATTO488 PPE-recht in levende celmembranen (A) Schematische weergave van ATTO488 PPE-insertie in celmembraan (B):.. Een ROI (rode box) wordt geselecteerd in een grotendeels uniform deel van de cel, het vermijden van cel grens en zeer fluorescerende vlekken. (C) De diffusie wet gemeten in het geselecteerde ROI toont een platte gedrag bevestigt een vrije diffusie model voor deze component. Klik hier om een grotere versie te bekijken dit cijfer.

Figuur 5 TfR-GFP diffusie wet levende celmembranen (A) Schematische weergave van TfR-GFP insertie in celmembraan. De cytoplasmatische staart van de receptor interageert met het membraan skelet, die als omheining voor receptor mobiliteit (B. ) TIRF beeld van CHO-GFP expressie TfR: a ROI is geselecteerd voorkeur lage expressie brengen artefacten wegens overexpressie voorkomen (C) De diffusie wet van TfR (zwarte stippen), anders PPE (grijze lijn uit figuur 4). toont het typische gedrag van gedeeltelijke afsluiting diffusie waarbij een eerste vlakke gedeelte wordt gevolgd door een daling van de D app. Klik hier om een grotere versie van deze fi bekijkenguur.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Single Particle Tracking (SPT) is een van de meest voorkomende strategieën om moleculaire dynamica te bestuderen en het heeft het grote voordeel van het meten van deeltjestrajecten. Hierdoor kan sonderen het gedrag van zelfs enkele gemerkte deeltjes in een complex systeem. Echter, dit voordeel SPT heeft typisch een lage dichtheid van de probe en zeer helder labels bereikt. In dit geval een ingewikkelde procedure productie, etikettering en insertie in het systeem nodig: bijzonder hoge temporele resolutie (usec range) anorganische probe zijn gewoonlijk vereist (bijvoorbeeld quantum dots of metaal nanodeeltjes) krijgen. Vergeleken met SPT de huidige werkwijze toont een aantal belangrijke voordelen. Ten eerste kan deze benadering worden gebruikt in conjugatie met fluorescente proteïnen. Aldus, vergeleken met SPT, is een hogere temporele resolutie bereikt (op hetzelfde label) door het lagere aantal fotonen vereist 34. Meer in detail, deze eigenschap maakt het indrukken van de temporale resolution hieronder 10-3 sec ook bij gebruik van codeerbare fluorescerende eiwitten, en deze termijn geeft exclusieve toegang tot de nanoschaal dynamiek van membraan bestanddelen. Tenslotte zij opgemerkt dat moleculaire diffusie wetten beschreven door analyse volledige ruimtetijd correlatiefunctie, zonder de noodzaak om elk molecuul te volgen.
De vergelijking met STED-gebaseerde FCS is ook interessant. In een STED FCS-meting van de gemiddelde looptijd van moleculen gedurende verlagen waarneming volumes gemeten door de temporele correlatie van het fluorescentiesignaal. Dit maakt het verkrijgen van een lokale meting van de moleculaire dynamica ook onder de diffractie limiet. In de gepresenteerde aanpak de diffusie wet wordt gemeten als het gemiddelde van alle deeltjes bewegen in de geselecteerde ROI waargenomen door standaard diffractie-beperkt observatie volume. Echter, gerapporteerde resultaten tonen aan dat deze werkwijze niet beperkt door diffractie, maar alleen the temporele resolutie beschikbaar. In feite, hoewel een buigingsbegrensde overname wordt gebruikt om fluctuaties (analoog met wat gebeurt in andere super-resolutie technieken zoals PALM en STORM), moleculaire verplaatsingen ruim onder de diffractie worden opgespoord kan worden (direct) berekend, zoals reeds aangetoond met behulp PEN meten moleculaire stromen 32. Bovendien, in tegenstelling STED-FCS, deze benadering kan eenvoudig worden toegepast op een breed scala aan commerciële en bestaande microscopie opstellingen, zoals raster scanning microscopen of groothoek-camera gebaseerde microscopen. Het is het vermelden waard dat STED-FCS metingen van moleculaire diffusie wetten strikt behoeven een fluorofoor-afhankelijke kalibratie van de grootte van de instrumentale taille. Tegengesteld, de meting gepresenteerd hier niet een kalibratie van het systeem (alleen nodig voor de schatting van deeltjesgrootte) vereisen.
De werkelijke resolutie in de meting van de deeltjes verplaatsingen door de gepresenteerde methode deaf van hoe nauwkeurig we de correlatiefunctie kan meten. Derhalve wordt het niet inherent beperkt door diffractie, analoog aan het geval SPT dat de resolutie afhankelijk van de nauwkeurigheid waarmee het partikel "beeld" gemeten. Een significante correlatie in minder dan 1 min meten van de voorgestelde experimenten weinig fotonen (gewoonlijk onder 10 fotonen) voor elk deeltje in elk frame genoeg. In feite, is de bijdrage van alle waargenomen deeltjes samen gemiddeld wanneer de correlatie-functie wordt berekend, ook als deeltjes niet zijn geïsoleerd. Deze eigenschap is inherent van fluctuatie correlatie methoden en maakt gebruik van dim en dichte labels, zoals fluorescerende eiwitten getransfecteerd in levende cellen.
Daarom lijkt het duidelijk dat de minimale meetbare verplaatsing is afhankelijk van de diffusie van de deeltjes en de tijdsresolutie van het beeldvormende opstart. Als voorbeeld, overweeg diffusie van moleculen op de celmembraan,waarbij de maximaal gemeten diffusie van eiwitten of lipiden ongeveer 5 urn 2 sec -1. Onder dergelijke omstandigheden is een tijdsresolutie van ongeveer 10 -4 sec moeten we een gemiddelde verplaatsing van 50 nm vangen. Deze tijdresolutie kan door snelle scanning microscopen langs enkelvoudige verbindingen of door snelle EMCCD camera, waar tijdresolutie samenvalt met de belichtingstijd, zoals ze vertoonden.
Een bijkomende essentiële eis voor deze methode om moleculaire dynamica nauwkeurig te beschrijven een juiste ruimtelijke verdeling. In feite, om de correlatiefunctie passen we een ruimtelijke verdeling (pixelgrootte) hoger dan de taille van de instrumentele PSF. In de meeste commerciële microscopen (confocale of breed), PSF taille reikt van 200 nm tot 500 nm (hoofdzakelijk afhankelijk van de numerieke opening van het geselecteerde doel en de golflengte) en kan gemakkelijk worden gemeten met een kalibratie experiment met nano sized fluorescerende kralen. THus een pixelgrootte van 70-150 nm (3 keer hoger dan de instrumentele middel) kan volstaan. Echter, de pixelgrootte worden aangepast aan het bestudeerde systeem rekening houdend met een eenvoudige regel: verlaag de pixelgrootte groter de nauwkeurigheid in de beschrijving van de correlatiefunctie. Bovendien is de minimale grootte van het beeld te verwerven moet minstens 3 keer groter is dan de maximale verplaatsing van belang (plus de instrumentale taille) zijn. Dit is nodig om een goede convergentie van de fitting algoritme en een statistisch significante steekproef van moleculaire verplaatsingen bereikt. Als voorbeeld het molecuulgewicht verplaatsingen kleiner dan enkele honderden nanometers (bijvoorbeeld 200 nm) beeldformaat enkele microns voldoende bestuderen. Bovendien is het totale aantal pixels (waarbij constant de pixelgrootte) effecten op de kwaliteit van de correlatiefunctie. In feite, maakt het mogelijk een grotere afbeelding van gemiddeld meer informatie in de correlatie-functie, zelfs als ten koste van de tijd resolution. Met betrekking tot de camera-gebaseerde systeem hier gebruikt, dan kunt u er rekening mee dat de fysieke grootte van de pixel op de chip is bevestigd. Dientengevolge verminderen de pixelgrootte verlaagt het signaal in elke pixel (die afhangt van de kwadraat van de pixelgrootte), vermindert het gezichtsveld, en vereist hogere vergrotingsfactor. Anderzijds, in een scansysteem, waarbij het observatiegebied is vastgesteld verminderen van de pixelgrootte resulteert gewoonlijk in een groter aantal pixels ten koste van de tijdsresolutie.
Weinig details over de detector gebruikt moeten worden besproken. Unlike single-photon detectoren, EMCCD meting van een gemiddelde intensiteit (digitaal niveau, DL) die niet recht evenredig met de verzamelde licht door de aanwezigheid van een offset. Hoewel deze offset lager is dan de dynamische bereik van de camera (enkele honderden tegenover 2 16 in 16 bits aflezing) en verwaarloosbaar experimenten waarbij vele fotonen worden verzameld, moet in aanmerking worden genomenverkrijgen van een correcte normalisatie van de correlatiefunctie. Ook de offset kan gebruikt worden als referentie bij weinig licht om de hoeveelheid signaal verzameld identificeren. Bovendien, teneinde de gemiddelde hoeveelheid fotonen die worden verzameld tijdens de verwerving schatten de gemiddelde digitaal niveau gekoppeld aan elke foton verzameld worden gemeten. Deze hoeveelheid kan worden opgehaald door het blootstellen van de camera aan een zeer lage lichtintensiteit (bijvoorbeeld het achtergrondlicht in de kamer); in feite, in dit geval, we redelijkerwijs aannemen dat slechts enkele fotonen bereikt de camera, dat wil zeggen de gemeten intensiteit kan worden gerelateerd aan slechts nul of een foton.
Tenslotte, laten we spreken over hoe sommige alternatieve meetsystemen (bv verschillende opstellingen microscopie) kan worden gebruikt om de gepresenteerde metingen. Allereerst kan het "W'factor in Vergelijking 2 (dat staat voor de autocorrelatie van de instrumentele PSF) worden aangepast thij bijzonder meetsysteem gebruikt om de experimentele correlatiefunctie passen. Zoals eerder aangetoond 34, een eenvoudige zaak de verwerving whit een laser scanning microscoop wanneer de scansnelheid aanzienlijk hoger dan deeltje dynamiek. In een dergelijk geval, in feite, de beweging van deeltjes tijdens de acquisitie tijd (dwz lijntijd) is te verwaarlozen en de correlatiefunctie goed benaderd door een Gaussische functie. In de context van de nieuwe beeldvormende technieken, wordt een interessante benadering gebaseerd op de mogelijkheid om zeer dunne lichte platen (1-2 um) geproduceerd door het monster 41. De lichte mogelijkheid biedt selectieve belichting van een enkel vlak (Single vliegtuig verlichting microscopie SPIM) in het monster en, gecombineerd met een camera gebaseerde acquisitiesysteem snel optische sectie in 3D 42. Vanwege deze kenmerken, is SPIM succes geconjugeerd met FCS 43 en kon een vali vertegenwoordigend hulpmiddel om de gepresenteerde analyse te breiden tot 3D omgevingen zoals het cytoplasma of de nucleus van levende cellen.
Samenvattend, vanuit experimenteel standpunt deze benadering is alleen de toegang tot een microscoop met een snelle acquisitie module. Het eiwit van belang kan worden gelabeld met fluorescerende eiwit of organisch fluorofoor, dus ook waardoor multicolor imaging. In deze context, ogen we de mogelijkheid cross gebruik i MSD analyse subpopulaties moleculen selecteren en onthullen interacties en co-diffusie van levende celmembranen. Tot slot zijn wij van mening dat deze aanpak een krachtig hulpmiddel om eiwitten en / of vetten ondergaan dynamische partitionering binnen nanodomains op het plasmamembraan te bestuderen kunnen vertegenwoordigen. In dit geval, de zeer uiteenlopende grootte en levensduur van de nanodomains introduceren extra complexiteit in reële gegevens die verder methoden implementaties zijn 2-kleuren weergave vereist, lokaleanalyse (bijvoorbeeld 2D pair correlatie) en / of fluorescentie anisotropie.
Subscription Required. Please recommend JoVE to your librarian.
Materials
| Name | Company | Catalog Number | Comments |
| iXon Ultra 897 | Andor | DU-897U-CS0 | |
| Solis | Andor | ||
| CHO-K1 | ATCC | CCL-61 | |
| ATTO 488 labeled PPE | ATTO-TEC GmbH | AD 488-151 | |
| DOPE | Avanti Polar Lipids, Inc. | 850725 | |
| DOTAP | Avanti Polar Lipids, Inc. | 890890 | |
| 100x Penicillin-Streptomycin-Glutamine | Gibco | 10378-016 | |
| DMEM/F-12 | Gibco | 21331 | |
| FBS | Gibco | 10082147 | |
| HEPES | Gibco | 15630-106 | |
| PBS | Gibco | 10010-023 | |
| SimFCS 3.0 | Globals Software | the software can be downloaded here: http://www.lfd.uci.edu/globals/ | |
| DMI6000 with TIRF modulus | Leica | ||
| LAS AF | Leica | ||
| Lipofectamine 2000 | Lipofectamine | 11668019 | |
| Matlab | MathWork | ||
| ImageJ | NIH |
| Name | Company | Catalog Number | Comments |
| C-terminal GFP tagged Tranferrin Receptor | OriGene | RG200980 | |
| Agar | Sigma Aldrich | A5306 | |
| Chloroform | Sigma Aldrich | 528730 | |
| Latex beads, fluorescent yellow-green, 30 nm | Sigma Aldrich | L5155 | |
| SONICA Ultrasonic Cleaners | SOLTEC | ETH S3 | |
| Petri Dishes | Willco | GWSt-3522 | |
| Bio-Format importer for Matlab | http://www.openmicroscopy.org/site/support/bio-formats5/users/matlab/ | ||
| ICS-MatLab Tools | https://www.cellmigration.org/resource/imaging/software/ICSMATLAB_28-02-06.zip | ||
| Simulation by Matlab Tutorial | https://www.cellmigration.org/resource/imaging/icsmatlab/ICSTutorial.html | ||
| Simulation by SimFCS Tutorial | https://www.cellmigration.org/resource/imaging/ppt-pdf/RICS%20Simulations.ppt |
References
- Engelman, D. M. Membranes are more mosaic than fluid. Nature. 438 (7068), 578-580 (2005).
- Vereb, G., et al. yet structured: The cell membrane three decades after the Singer-Nicolson model. Proc. Natl. Acad. Sci. U. S. A. 100 (14), 8053-8058 (1073).
- Ishihara, A., Hou, Y., Jacobson, K. The Thy-1 antigen exhibits rapid lateral diffusion in the plasma membrane of rodent lymphoid cells and fibroblasts. 84 (5), 1290-1293 (1987).
- Axelrod, D., et al. Lateral motion of fluorescently labeled acetylcholine receptors in membranes of developing muscle fibers. Proc. Natl. Acad. Sci. U. S. A. 73 (12), 4594-4598 (1976).
- Jacobson, K., Derzko, Z., Wu, E. S., Hou, Y., Poste, G. Measurement of the lateral mobility of cell surface components in single, living cells by fluorescence recovery after photobleaching. J. Supramol. Struct. 5 (4), 10-1002 (1976).
- Kusumi, A., et al. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annu. Rev. Biophys. Biomol. Struct. 34, 351-378 (2005).
- Kusumi, A., Ike, H., Nakada, C., Murase, K., Fujiwara, T. Single-molecule tracking of membrane molecules: plasma membrane compartmentalization and dynamic assembly of raft-philic signaling molecules. Semin. Immunol. 17 (1), 3-21 (2005).
- Schwille, P., Korlach, J., Webb, W. W. Fluorescence correlation spectroscopy with single-molecule sensitivity on cell and model membranes. Cytometry. 36, 176-182 (1999).
- Gielen, E., et al. Diffusion of sphingomyelin and myelin oligodendrocyte glycoprotein in the membrane of OLN-93 oligodendroglial cells studied by fluorescence correlation spectroscopy. C. R. Biol. 328 (12), 1057-1064 (2005).
- Weiss, M., Hashimoto, H., Nilsson, T. Anomalous protein diffusion in living cells as seen by fluorescence correlation spectroscopy. Biophys. J. 84, 4043-4052 (2003).
- Wawrezinieck, L., Rigneault, H., Marguet, D., Lenne, P. F. Fluorescence correlation spectroscopy diffusion laws to probe the submicron cell membrane organization. Biophys. J. 89 (6), 4029-4042 (2005).
- Lenne, P. F., et al. Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork. EMBO J. 25 (14), 3245-3256 (2006).
- Ries, J., Schwille, P. Studying slow membrane dynamics with continuous wave scanning fluorescence correlation spectroscopy. Biophys. J. 91 (5), 1915-1924 (2006).
- Ruan, Q., Cheng, M. A., Levi, M., Gratton, E., Mantulin, W. W. Spatial-temporal studies of membrane dynamics: scanning fluorescence correlation spectroscopy (SFCS). Biophys. J. 87 (2), 1260-1267 (2004).
- Berland, K. M., So, P. T., Chen, Y., Mantulin, W. W., Gratton, E. Scanning two-photon fluctuation correlation spectroscopy: particle counting measurements for detection of molecular aggregation. Biophys. J. 71, 410-420 (1996).
- Heinemann, F., Betaneli, V., Thomas, F. A., Schwille, P. Quantifying lipid diffusion by fluorescence correlation spectroscopy: a critical treatise. Langmuir. 28 (37), 13395-13404 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Capturing directed molecular motion in the nuclear pore complex of live cells. Proc. Natl. Acad. Sci. U. S. A. 109 (25), 9863-9868 (2012).
- Sanchez, S. A., Tricerri, M. A., Gratton, E. Laurdan generalized polarization fluctuations measures membrane packing micro-heterogeneity in vivo. Proc. Natl. Acad. Sci. U. S. A. 109 (19), 7314-7319 (2012).
- Cardarelli, F., Lanzano, L., Gratton, E. Fluorescence correlation spectroscopy of intact nuclear pore complexes. Biophys. J. 101 (4), 27-29 (2012).
- Di Rienzo, C., et al. Unveiling LOX-1 receptor interplay with nanotopography: mechanotransduction and atherosclerosis onset. Sci. Rep. 3, 10-1038 (2013).
- Unruh, J. R., Gratton, E. Analysis of molecular concentration and brightness from fluorescence fluctuation data with an electron multiplied CCD camera. Biophys. J. 95 (11), 5385-5398 (2008).
- Kannan, B., et al. Electron multiplying charge-coupled device camera based fluorescence correlation spectroscopy. Anal. Chem. 78 (10), 3444-3451 (2006).
- Jones, S. A., Shim, S. H., He, J., Fast Zhuang, X. three-dimensional super-resolution imaging of live cells. Nat. Methods. 8 (6), 499-508 (2011).
- Rust, M. J., Bates, M., Zhuang, X. Sub-diffraction-limit imaging by stochastic optical reconstruction microscopy. 3 (10), 793-795 (2006).
- Betzig, E., et al. Imaging intracellular fluorescent proteins at nanometer resolution. Science. 313 (5793), 1642-1645 (2006).
- Hess, S. T., Girirajan, T. P., Mason, M. D. Ultra-high resolution imaging by fluorescence photoactivation localization microscopy. Biophys. J. 91 (11), 4258-4272 (2006).
- Manley, S., et al. High-density mapping of single-molecule trajectories with photoactivated localization microscopy. Nat. Methods. 5 (2), 155-157 (2008).
- Hell, S. W. Far-field optical nanoscopy. Science. 316 (5828), 1153-1158 (2007).
- Klar, T. A., Hell, S. W. Subdiffraction resolution in far-field fluorescence microscopy. Opt. Lett. 24 (14), 954-956 (1999).
- Eggeling, C., et al. Direct observation of the nanoscale dynamics of membrane lipids in a living cell. Nature. 457 (7233), 1159-1162 (2009).
- Hedde, P. N., et al. Stimulated emission depletion-based raster image correlation spectroscopy reveals biomolecular dynamics in live cells. Nat. Commun. 4, Forthcoming.
- Hebert, B., Costantino, S., Wiseman, P. W. Spatiotemporal image correlation spectroscopy (STICS) theory, verification, and application to protein velocity mapping in living CHO cells. Biophys. J. 88 (5), 3601-3614 (2005).
- Brown, C. M., et al. Probing the integrin-actin linkage using high-resolution protein velocity mapping. J. Cell Sci. 119, 5204-5214 (2006).
- Di Rienzo, C., Gratton, E., Beltram, F., Cardarelli, F. Fast spatiotemporal correlation spectroscopy to determine protein lateral diffusion laws in live cell membranes. Proc. Natl. Acad. Sci. U. S. A. 110 (30), 12307-12312 (2013).
- Mueller, V., et al. STED nanoscopy reveals molecular details of cholesterol- and cytoskeleton-modulated lipid interactions in living cells. Biophys. J. 101 (7), 1651-1660 (2011).
- Kleusch, C., Hersch, N., Hoffmann, B., Merkel, R., Csiszar, A. Fluorescent lipids: functional parts of fusogenic liposomes and tools for cell membrane labeling and visualization. Molecules. 17 (1), 1055-1073 (2012).
- Ries, J., Chiantia, S., Schwille, P. Accurate determination of membrane dynamics with line-scan FCS. Biophys. J. 96 (5), 1999-2008 (2009).
- Kolin, D. L., Wiseman, P. W. Advances in image correlation spectroscopy: measuring number densities, aggregation states, and dynamics of fluorescently labeled macromolecules in cells. Cell Biochem. Biophys. 49 (3), 141-164 (2007).
- Digman, M. A., et al. Measuring fast dynamics in solutions and cells with a laser scanning microscope. Biophys. J. 89 (2), 1317-1327 (2005).
- Ritchie, K., et al. Detection of non-Brownian diffusion in the cell membrane in single molecule tracking. Biophys. J. 88 (3), 2266-2277 (2005).
- Voie, A. H., Burns, D. H., Spelman, F. A. Orthogonal-plane fluorescence optical sectioning: three-dimensional imaging of macroscopic biological specimens. J. Microsc. 170, 229-236 (1993).
- Huisken, J., Swoger, J., Del Bene,, Wittbrodt, F., J,, Stelzer, E. H. Optical sectioning deep inside live embryos by selective plane illumination microscopy. Science. 305 (5686), 1007-1009 (2004).
- Wohland, T., Shi, X., Sankaran, J., Stelzer, E. H. Single plane illumination fluorescence correlation spectroscopy (SPIM-FCS) probes inhomogeneous three-dimensional environments. Opt. Express. 18 (10), 10627-10641 (2010).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}