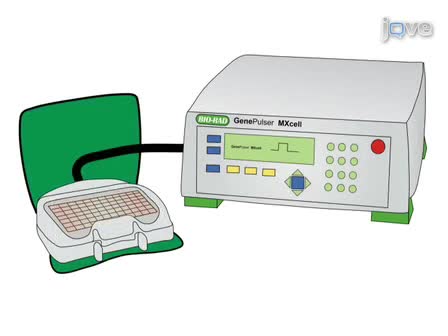
This manuscript describes a novel technique which allows the detection of intracellular gene expression after endocytosis of fluorescence-labeled nanoparticles directly in live cells. The method does not require manipulation of the cells and is not restricted with respect to the target gene or species.