


Summary
Aktomiyozin demetlerinin in vitro oluşumu ve optik cımbız kullanılarak miyozin topluluk kuvveti oluşumunun ölçülmesi sunulmuş ve tartışılmıştır.
Abstract
Miyosinler, ATP'yi aktin filament (AF) izleri boyunca adım atmak için hidrolize eden motor proteinlerdir ve motilitesi ve kas kasılması gibi hücresel süreçlerde gereklidir. Kuvvet üretme mekanizmalarını anlamak için, miyozin II hem tek moleküllü (SM) seviyede hem de optik yakalama gibi biyofiziksel yöntemler kullanılarak in vitro motor ekipleri olarak araştırılmıştır.
Bu çalışmalar, üç boncuklu bir düzenlemedeki tek moleküllü seviyeden, sert bir boncuk veya kayma düzeninde örtü kayması yüzeyinde birlikte çalışan motor gruplarına geçerken miyozin kuvvet üretme davranışının büyük ölçüde farklılık gösterebileceğini göstermiştir. Bununla birlikte, bu tahlil yapıları, miyozin grup dinamiklerinin viskoelastik yapısal hiyerarşi içinde bir hücre içinde olduğu gibi değerlendirilmesine izin vermez. Çoklu aktin filamentlerle etkileşime giren miyozin toplulukları tarafından kuvvet oluşturma mekaniğini araştırmak için optik cımbız kullanan bir yöntem geliştirdik.
Bu aktomiyozin demetleri, motor iletişimi ve topluluk kuvveti çıkışını yakalayan hiyerarşik ve uyumlu bir ortamda araştırmayı kolaylaştırır. Tahlilin özelleştirilebilir doğası, miyozin topluluğundaki, aktin filament demetindeki veya çevredeki ortamdaki değişikliklerin farklı kuvvet çıktılarına nasıl yol açtığını anlamak için deneysel koşulların değiştirilmesine izin verir.
Introduction
Motor proteinleri, kimyasal enerjiyi mekanik çalışmaya dönüştürerek yaşam için gereklidir 1,2,3. Myosin motorları, bir ize benzer filamentler boyunca adımlar atarak aktin filamentleri ile etkileşime girer ve aktin-miyozin ağlarının dinamikleri, diğer temel görevlerin yanı sıra kas kasılması, hücre hareketliliği, sitokinezi sırasında kasılma halkası ve hücre içindeki yükün hareketini gerçekleştirir 3,4,5,6,7,8 . Miyosinlerin çok önemli rolleri olduğundan, miyozin-aktin ağının işlevselliğindeki başarısızlık, hipertrofik kardiyomiyopatide (HCM) kalp hiperkontraktilitesine neden olan miyozin ağır zincirindeki mutasyonlar gibi hastalık gelişimine yol açabilir 9,10,11,12,13,14 . Kas kasılmasında, bireysel miyozin motorları, AF'lerin 4,15,16,17,18'in göreceli kaymasını gerçekleştiren gerekli mekanik enerjiyi sağlamak için bir topluluk olarak çalışarak birbirleriyle işbirliği yapar. Myosin motorları, AF'ler arasında çapraz köprüler oluşturur ve hizalanmış filamentlerin17,18,19,20,21 dikenli ucuna doğru toplu olarak hareket etmek için mekanokimyasal döngüsü nedeniyle konformasyonel değişiklikler kullanır.
Optik yakalama gibi teknikler kullanılarak SM düzeyinde kantitatif in vitro motiliteli testlerin geliştirilmesi, SM kuvvet üretiminin ve adım boyutlarının ölçülmesi de dahil olmak üzere bireysel miyozin motorlarının nasıl çalıştığına dair benzeri görülmemiş ayrıntıların toplanmasını kolaylaştırmıştır 22,23,24,25,26,27,28,29,30 . Finer ve ark., tek miyozin II motorlarının kuvvet oluşturma mekaniğini araştırmak için "üç boncuk" veya "dambıl" optik yakalama testini geliştirdiler23,31. Kas miyozin II, AF'leri sözleşmek için ekipler halinde çalıştığından, ancak SM seviyesinde işlemsel olmadığından, optik yakalama testi oryantasyonunun klasik motora bağlı boncuk yaklaşımı32'den yeniden düzenlenmesi gerekiyordu. Dambıl tahlilini oluşturmak için, bir AF'yi bir kapaklı boncuğa bağlı bir miyozin motoru üzerinde tutmak için iki optik tuzak kullanıldı ve tek motor tarafından kuvvet çıkışı, AF'nin tuzak23 içindeki hareketleriyle ölçüldü.
Bununla birlikte, SM kuvvetleri ve tek bir motor / tek filament tahlili oryantasyonu kullanarak, sistem düzeyinde kuvvet üretimi hakkında tam bir görüntü vermez, çünkü miyozin II de dahil olmak üzere birçok motor proteini izolasyonda çalışmaz ve genellikle parçalarının toplamı olarak işlev görmez 15,16,17,32,33,34,35,36 . Birden fazla filament ile etkileşime giren birden fazla motor içeren daha karmaşık yapılar, miyozin ve aktin filamentlerinin ağlarının sinerjisini daha iyi anlamak için gereklidir15,32. Dambıl tahlili oryantasyonu, bir boncuğa bağlı birden fazla miyozin bulundurarak veya bir yüzeye bağlı miyozin kalınlığında bir filament kullanarak ve motorların asılı AF 4,23,34,37,38,39,40 ile etkileşime girmesine izin vererek küçük topluluk kuvveti oluşumunu araştırmak için kullanılmıştır.
Diğer küçük topluluk tahlilleri arasında, miyozin motorlarının bir kapak kayması yüzeyine kaplandığı ve 4,35,36,38,39,40,41,42,43 motor ekibi tarafından üretilen kuvveti araştırmak için bir AF'ye bağlı bir boncuk kullanıldığı bir in vitro filament kayma testi bulunmaktadır. . Her iki durumda da, miyosinler sert bir yüzeye bağlanır - boncuk veya örtü kayması - ve bir AF kullanır. Bu gibi durumlarda, motorlar serbestçe hareket edemez veya birbirleriyle iletişim kuramazlar ve miyozinlerin katı bir şekilde bağlanmış olması, motorların sarkomer32'de birlikte çalışacağı uyumlu, hiyerarşik ortamı yansıtmaz. Önceki çalışmalar, miyozin II'nin çevresini algılayabildiğini ve kuvvet üretimi ve görev oranı41,44,45 gibi özellikleri değiştirerek değişen viskoelastik veya motor konsantrasyon koşullarına göre uyum sağlayabileceğini öne sürmüştür. Bu nedenle, miyozin II topluluk kuvveti üretiminin mekanik temellerinin daha gerçekçi bir resmini çizmek için motor iletişimini ve sistem uyumluluğunu teşvik eden ve yakalayan bir optik yakalama testi geliştirmeye ihtiyaç vardır.
Burada, iki aktin filamenti arasında etkileşime giren çoklu miyozin motorlarından oluşan aktomiyozin demetleri veya sandviçler oluşturarak hiyerarşik yapıyı in vitro optik yakalama ile birleştirmek için bir yöntem geliştirdik. Bu modüler tahlil geometrisi, moleküler ve çevresel faktörlerin topluluk miyozin kuvveti üretimini nasıl etkilediğini doğrudan araştırma yeteneğine sahiptir. Ayrıca, bu aktin-miyozin toplulukları aracılığıyla kuvvet oluşturma mekanizmalarının araştırılması, kas kasılması gibi büyük ölçekli hücresel görevlerinmoleküler seviye 9,10,13'ten nasıl yayıldığını modellemeye ve anlamaya yardımcı olma potansiyeline sahiptir.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Gravür kapakları
- 100 g KOH'u 300 mL'lik %100 etanol içinde 1.000 mL'lik bir beherde eritin. KOH'un çoğunluğu çözülene kadar bir karıştırma çubuğuyla karıştırın.
DİKKAT: Konsantre KOH çözeltisi yanıklara ve giysilerde hasara neden olabilir. Eldiven, göz koruması ve laboratuvar önlüğü giyin. - Kapak fişlerini tek tek coverslip temizleme raflarına yerleştirin.
NOT: Raflar, kapak kaymasının her bir yüzünde aşındırma ve durulamaya, alttaki delikleri boşaltmaya ve sert aşındırma koşullarına dayanabilecek malzemeden yapılmasına izin vermek için aralıklı tek kapak kaymalarını tutan yarıklarla tasarlanmıştır. Ismarlama yapılabilir veya ticari olarak satın alınabilirler. - Üç adet 1.000 mL beher hazırlayın ve etiketleyin: biri 300 mL etanol ve iki tane 300 mL ters ozmoz (RO) suyu ile.
NOT: Burada, RO suyu bir laboratuvar su arıtma cihazından elde edilmiştir, ancak yerel bir arıtma cihazı mevcut değilse ticari olarak da satın alınabilir. - Dört beherin her birini 5 dakika boyunca gaz çıkarmak için bir banyo sonikatörüne yerleştirin.
- KOH ve etanol beherine bir kapak kapağı rafını batırın ve 5 dakika boyunca sonikasyon yapın.
- Kapak kapaklarının rafını KOH/etanol beherinden sadece etanol içeren behere aktarın. Boncuklanma kalmayana kadar beherin içine yukarı ve aşağı daldırın.
NOT: Kapak kapaklarını rahatsız etmemeye veya rafı zorla beherin içine düşürmemeye dikkat edin. Bu, kapak kaymalarının raftan çıkmasına veya kimyasal sıçramaya neden olur. - Kapak kapaklarının rafını dikkatlice etanol beherinden bir su kabına aktarın, boncuk kalmayana kadar yukarı ve aşağı daldırın.
- Kapak kapaklarının rafını henüz kullanılmamış su kabına batırın ve 5 dakika boyunca tekrar sonikleştirin.
- Kapak kapakları rafını, kapaklardan düzgün bir şekilde akana kadar suyla püskürtmek için bir şişe kullanın. Etanol ile tekrarlayın.
- Rafları 20 dakika boyunca 90 °C'de bir fırında kurumaya bırakın. Kazınmış kapak kapaklarının raflarını, kullanımdan önce kontaminasyonu önlemek için oda sıcaklığında kapalı kaplarda saklayın.
2. Aktin filament polimerizasyonu
- Çözüm T Yapın
- 50 mL'lik bir konik tüpte, 3.94 g Tris-HCl ve 0.147 g CaCl2 ekleyin. Toplam 50 mL hacim yapmak için RO suyu ekleyin ve iyice karıştırın.
NOT: Çözelti T'nin nihai konsantrasyonları sırasıyla 500 mM Tris-HCl ve 20 mM CaCl2'dir . - Tüp Solution T'yi etiketleyin ve 4 ° C'de saklayın.
- 50 mL'lik bir konik tüpte, 3.94 g Tris-HCl ve 0.147 g CaCl2 ekleyin. Toplam 50 mL hacim yapmak için RO suyu ekleyin ve iyice karıştırın.
- TC Arabelleği Yapın
- 50 mL'lik konik bir tüpte 40 mL RO suyu ve 1,5 mL Çözelti T'yi karıştırın. Az miktarda konsantre KOH ekleyerek pH'ı 8.0'a değiştirin. Çözeltinin 50 mL'sini yapmak için su ekleyin ve pH'ı doğrulayın. Gerekirse pH'ı ayarlayın.
NOT: Son TC tamponu, pH 8'de 5 mM Tris-HCl ve 0,2 mM CaCl2 içerir. - TC tüpünü etiketleyin ve 4 °C'de saklayın.
- 50 mL'lik konik bir tüpte 40 mL RO suyu ve 1,5 mL Çözelti T'yi karıştırın. Az miktarda konsantre KOH ekleyerek pH'ı 8.0'a değiştirin. Çözeltinin 50 mL'sini yapmak için su ekleyin ve pH'ı doğrulayın. Gerekirse pH'ı ayarlayın.
- FC Arabelleği Yap
- 100 mL'lik bir tampon şişeye 85 mL RO suyu, 10 mL Çözelti T, 3.73 g KCl ve 0.041 g MgCl2 ekleyin. Küçük miktarlarda konsantre KOH ekleyerek pH'ı 7.5 olarak değiştirin. Son hacmi 100 mL'ye çıkarmak için su ekleyin ve pH'ı doğrulayın.
NOT: Son FC tamponu, pH 7,5'te 500 mM Tris-HCl, 500 mM KCl, 2 mM MgCl2 ve 2 mM CaCl2 içerir. - FC tüpünü etiketleyin ve 4 °C'de saklayın.
- 100 mL'lik bir tampon şişeye 85 mL RO suyu, 10 mL Çözelti T, 3.73 g KCl ve 0.041 g MgCl2 ekleyin. Küçük miktarlarda konsantre KOH ekleyerek pH'ı 7.5 olarak değiştirin. Son hacmi 100 mL'ye çıkarmak için su ekleyin ve pH'ı doğrulayın.
- Genel Aktin Arabelleğini (GAB) hazırlayın.
- Bir mikrosantrifüj tüpünde 485 μL TC tamponu, 10 μL 10 mM ATP ve 5 μL 50 mM DTT karıştırın.
NOT: Son tampon koşulları 5 mM Tris-HCl, 0,2 mM CaCl 2, 0,5 mM DTT ve0,2 mM ATP'dir. - GAB olarak etiketleyin ve 4 °C'de saklayın.
- Bir mikrosantrifüj tüpünde 485 μL TC tamponu, 10 μL 10 mM ATP ve 5 μL 50 mM DTT karıştırın.
- Aktin Polimerizasyon Tamponunu (APB) hazırlayın.
- Bir mikrosantrifüj tüpünde 455 μL FC tamponu, 25 μL 100 mM ATP ve 20 μL 50 mM DTT karıştırın.
NOT: Son tampon koşulları 50 mM Tris-HCl, 500 mM KCl, 2 mM MgCl 2,2 mM CaCl 22 mM DTT ve 5 mM ATP'dir. - Tüpü APB olarak etiketleyin ve 4 ° C'de saklayın.
- Bir mikrosantrifüj tüpünde 455 μL FC tamponu, 25 μL 100 mM ATP ve 20 μL 50 mM DTT karıştırın.
- Aktin'i yeniden oluşturmak
- 1 mg'lık bir liyofilize aktin şişesine 100 μL deiyonize su ekleyerek tavşan iskelet kası aktinini yeniden oluşturur. Hafifçe yukarı ve aşağı pipetleyerek, iyice karıştırın. Aliquot'u 5 μL numunelere dönüştürün, anında dondurun ve 10 mg / mL aktin alikotlarını -80 ° C'de saklayın.
- 20 μL RO suyu ekleyerek biyotinile tavşan iskelet kası aktinini yeniden oluşturur. Aliquot'u 5 μL numunelere koyun, anında dondurun ve 1 mg / mL biyotinile aktin alikotlarını -80 ° C'de saklayın.
- Rodamin falloidin stabilizasyonu ile etiketsiz aktin polimerizasyonu
- 10 mg / mL aktin'den bir şişe çözün ve buz üzerinde tutun.
- Taze GAB tamponu hazırlayın, aktin aliquot'a 100 μL GAB ekleyin ve hafifçe yukarı ve aşağı pipetleyerek, karıştırın. Çözeltiyi buz üzerinde 1 saat boyunca inkübe edin.
- Kuluçka sırasında taze APB hazırlayın. Kuluçkadan sonra, aktin çözeltisine 11 μL APB ekleyerek aktin'i filamentlere polimerize edin. Hafifçe yukarı ve aşağı pipetleyerek, iyice karıştırın. 20 dakika boyunca buzun üzerine yerleştirin.
- Taze polimerize aktin filament çözeltisine 5 μL rodamin etiketli falloidin ekleyin. Karanlıkta 1 saat boyunca buz üzerinde bırakın.
- Alüminyum folyoya sarılmış rodamin aktin şişesini karanlıkta 4 ° C'de saklayın.
NOT: Bu filamentlerin en fazla 1 hafta süreyle kullanılması önerilir. AF kalitesi, yalnızca AF'leri içeren bir akış hücresinin hızlı bir şekilde görüntülenmesi ve tutarlı filamentlerin günden güne görüntülenmesiyle her gün doğrulanabilir.
- Alexa Fluor 488 falloidin stabilizasyonu ile biyotinile aktin polimerizasyonu
- Bir şişe 10 mg / mL aktin ve 1 şişe 1 mg / mL biyotinillenmiş aktini çözün ve buz üzerinde tutun.
- Yeni GAB tamponu oluşturun.
- İki şişeyi (adım 2.8.1) 10: 1 aktinin: biyotinile aktin oranında birleştirin. Aktin karışımına 100 μL GAB ekleyin ve hafifçe yukarı ve aşağı pipetleyerek, iyice karıştırın. 1 saat boyunca buz üzerinde kuluçkaya yatırın.
- Kuluçka sırasında taze APB yapın.
- Kuluçka adımından sonra, aktin çözeltisine 11 μL APB ekleyerek aktin'i polimerize edin. Yavaşça yukarı ve aşağı pipetleyerek, iyice karıştırın. 20 dakika boyunca buz üzerinde kuluçkaya yatırın.
- 5 μL Alexa Fluor 488 etiketli falloidin ekleyin ve karanlıkta 1 saat boyunca buz üzerinde kuluçkaya yatırın.
- Alüminyum folyoya sarılmış biyotinile aktin şişesini karanlıkta 4 °C'de saklayın.
NOT: Bu filamentler maksimum 1 hafta süreyle kullanılabilir.
3. Myosin ve boncuk hazırlama
- Myosin II'yi yeniden oluşturma
- Standart bir minisantrifüj kullanarak tüpün dibinde toplamak için liyofilize iskelet miyozin II'yi kısaca (~ 5 s) aşağı doğru döndürün.
- RO suyunda hazırlanan 1 mM DTT'nin 100 μL'si eklenerek miyozini 10 mg/mL'ye yeniden oluşturun.
- RO suyunda 90 μL 1 mM DTT'ye 10 μL 10 mg / mL miyozin ekleyerek stok miyozin çözeltisini 10x seyreltin. Küçük hacimli (1-5 μL) alikotlar yapın, çıtçıtlı dondurma yapın ve -80 ° C'de saklayın.
NOT: Myosin aktivitesi, daha önce 46,47'de yayınlandığı gibi standart bir kayma filament testi yapılarak doğrulanabilir. Kısa bir açıklama için tartışmaya bakın.
- Streptavidin kaplı boncukların temizlenmesi
- 20 μL 1 μm streptavidin boncuklarını 80 μL RO suyuna seyreltin. 9.600 × g'da döndürerek ve 100 μL RO suyunda yeniden yapılandırarak dört kez yıkayın.
- % 40 genlikte 2 dakika boyunca sonikat yapın ve yıkanmış boncukları 4 ° C'de bir rotatörde saklayın.
4. Akış hücresi hazırlığı
- 50 mL'lik bir tüpe 30 mL% 100 etanol ekleyerek ve suya 200 μL% 0.1 w / v poli-l-lizin ekleyerek bir poli-l-lizin çözeltisi (PLL) hazırlayın ve iyice karıştırın.
- PLL çözeltisine kazınmış bir kapak kapağı ekleyin ve 15 dakika bekletin. Kapak kaymasını cımbızla çıkarın, tüpten yukarı çekilirken sadece kapak kaymasının kenarına dokunmaya dikkat edin (bkz. Şekil 1A-C). Kapakları eldivenli bir elinizle kenarlarından tutun.
- Kapak kapağını, etanol kalmayana ve kapak kapağında kalıntı kalmayana kadar filtrelenmiş bir havayolu ile kurulayın.
- Bir mikroskop slaytının ortasına, birbirinden 3-4 mm uzakta iki parça çift taraflı yapışkan bant uygulayın. Slaytın kenarından sarkan fazla bandı yırtın veya kesin.
- Bir kanal oluşturmak için mikroskop slaytının uzun eksenine dik olarak bandın üzerine PLL kaplı kapak kaymasını ekleyin (bir T oluşturarak).
- Kapak kaymasını bandın üzerine sıkıştırmak için küçük bir tüp kullanın ve mikroskop bant şeffaf olana kadar iyice kaydırın (Şekil 1A). Bantta kabarcık olmadığından emin olun, çünkü bu akış kanalından sızıntıya neden olabilir.
NOT: Akış hücresi 10-15 μL'lik bir hacmi tutabilir.
5. Aktomiyozin demeti hazırlama
- Ayrı tüplerde, saygın, etiketli aktininin 0,5 μL'sini 300 μL APB ile karıştırarak her bir aktin filament tipini (rodamin ve biyotinile 488 etiketli) 600x seyreltin. Her tüpe karşılık gelen etiketlenmiş falloidinden ilave 5 μL ekleyin ve karanlıkta 15 dakika boyunca buz üzerinde inkübe edin.
- Biyotinile aktin çözeltisine, 500 mg / mL'de 1 μL beta-D-glikoz, 25 mg / mL'de 1 μL glikoz oksidaz ve 500 birim / mL'de 1 μL katalaz oksijen süpürme sistemi ekleyin. 1 μL 100 mM ATP ve 1 μL 100x seyreltilmiş, temizlenmiş streptavidin boncukları ekleyin. Pipet ucu ile hafifçe karıştırın. Aktomiyozin demetinin geri kalanı monte edilirken süspansiyonu 4 ° C'de bir rotatöre koyun.
- PLL akış hücresine seyreltilmiş rodamin aktininin 15 μL'sini ekleyin (Şekil 1D). Fazla çözeltiyi akış hücresinden geçirin, ancak akış kanalının kurumasına izin vermeyin. Bir nem odasında 10 dakika boyunca kuluçkaya yatırın.
NOT: Nem odaları, ışığı engellemek için altına su eklenmiş boş pipet ucu kutularından ve alüminyum folyo kaplı kapaktan yapılabilir. - APB'de 1 mg/mL'lik bir kazein çözeltisi hazırlayın.
- Sonraki bileşenlerin spesifik olmayan bağlanmasını önlemek için 15 μL 1 mg / mL kazein ekleyin (Şekil 1E). Bir nem odasında 5 dakika boyunca inkübe edin.
- İstenilen miyozin konsantrasyonunu biyotinile aktin ve boncuk süspansiyonuna adım 5.2'den ekleyin. Pipet ucuyla hafifçe karıştırın ve ardından hemen akış hücresine 5.2 basamak süspansiyonunun 15 μL'sini + istenen miyozin konsantrasyonunu ekleyin (Şekil 1F, G). 20 dakika boyunca kuluçkaya yatırın. Görüntüleme ve optik yakalama deneyleri sırasında buharlaşmayı önlemek için akış hücresinin açık uçlarını oje ile kapatın.
NOT: 1 μM'lik bir miyozin çözeltisi konsantrasyonu sağlam bir paketleme sağlar ve tahlilin istenen özelleştirmesi için bir başlangıç noktası olarak kullanılabilir (bkz. Şekil 2).
6. Optik Tuzak (NT2 Nanotracker2) kullanarak kuvvet ölçümleri
NOT: Aşağıdaki protokol özellikle NT2 sistemi için olsa da, bu tahlil, floresan yeteneklerine sahip özel olarak üretilenler de dahil olmak üzere diğer optik yakalama sistemleriyle birlikte kullanılabilir. Genel iş akışı, slaytın yüzeyini odakta tutmak, boncuk kalibrasyonları yapmak ve floresan aktin demetlerini bularak veri elde etmekle aynı kalır. NT2 sistemi için, Ek Şekil S1, Ek Şekil S2, Ek Şekil S3, Ek Şekil S4, Ek Şekil S5, Ek Şekil S6 ve Ek Şekil S7, optik yakalama sistemi ve yazılım arabirimi hakkında ayrıntılı bilgi sağlar.
- Kontrol kutusunu ve lazeri açın (Ek Şekil S1).
- Masaüstündeki JPK Nanotracker simgesine tıklayarak optik tuzak bilgisayar yazılımını başlatın.
- Ortadaki Logitech düğmesine tıklayarak uzaktan kumandayı uyandırın (Ek Şekil S2).
- Açma/kapama düğmesini değiştirerek floresan modülünü açın (Ek Şekil S3).
- Parlak alan görüntüleme için filtre küpü taretini çevirin (Ek Şekil S4).
- Sistem hazır olduğunda, ekranın sol alt köşesindeki Lazer Gücü düğmesini kullanarak lazeri 50 mW'a kadar açın ve 30 dakika boyunca stabilize olmasına izin verin (Ek Şekil S5).
- Deney sırasında görüntüleme ve manipülasyon için bu pencereleri açmak üzere yazılım içindeki Aydınlatma, Kamera, Hedef ve Sahne Hareketi düğmelerine sırayla tıklayın. Açma/Kapama düğmesine tıklayarak mikroskop aydınlatmasını açın ve çubuğu tıklayıp sağa doğru sürükleyerek maksimum güce ayarlayın (Ek Şekil S5).
- Numune alanını açın ve numune tutucuyu mikroskop aşamasından çıkarın. Akış hücresini ekleyin, metal numune tutucularla sabitleyin ve kapak kaymalı kızağın altta olduğundan emin olun.
- Alt hedefin ortasına 30 μL RO suyu ekleyin. Pipet ucunun lense temas etmesine izin vermeyin. Örnek aşamayı yeniden ekleyin.
NOT: NT2 sistemi, yakalama hedefi olarak bir suya daldırma hedefi kullandığından, daldırma ortamı, kullanıcının kurulumundaki yakalama hedefine bağlı olarak farklı olabilir. - Su boncuğu kapak kaymasına dokunana kadar ekrandaki kontrol oklarını veya uzaktan kumandadaki L2'yi kullanarak alt hedefi yükseltin (Ek Şekil S5).
- Ekrandaki oklar veya uzaktan kumandadaki R2 kullanılarak akış hücresine olan mesafenin yaklaşık yarısına ulaşılana kadar üst hedefi düşürün. Akış hücresinin üst kısmına doğrudan üst hedefin altına 170 μL RO suyu ekleyin. Suyun yüzey gerilimini kırana ve bir menisküs oluşturana kadar üst hedefi alçaltın.
- Uzaktan kumanda üzerindeki ok tuş takımını kullanarak mikroskop sahnesini, akış kanalına bitişik bandın kenarına ulaşılana kadar hareket ettirin. Örnek kapıyı kapatın.
NOT: Numune kapağını kapattıktan sonra yapılan bir "tıklama", lazer deklanşörün artık açık olduğunu gösterir. Bu, yalnızca kapı kapalıysa deklanşörün açılmasına izin veren bir güvenlik özelliğidir. - Ekrandaki Hedef penceresini kullanarak, ekrandaki kontrolleri kullanarak üst oka tıklayarak Lazer Hedefi adlı alt hedefi yukarı getirerek bandın kenarını odaklayın. Alt oku tıklayarak üst hedef için de aynısını yapın (Ek Şekil S5).
NOT: Çift oklar hedefi veya sahne alanını daha hızlı hareket ettirir. Bandın kenarı odaklama için kullanılır, çünkü kapak kayması yüzeyine yakın büyük, bulunması kolay bir nesnedir. Bant içindeki hava kabarcıkları başka bir seçenektir. Ancak, kullanıcının yüzey odağını veya tercih edilen bir şirket içi yöntemi bulmak için otomatik bir rutini varsa bu gerekli değildir. - Bant netlendikten sonra, optik tuzağın üstündeki irisi kısmen kapatın. İrisin çokgen şekli görünene kadar üst hedefi aşağı indirin. Bu kenarları odaklayın, irisi yeniden açın ve ardından Asma kilit simgesine tıklayarak hedefleri birleştirin (Ek Şekil S5).
- Yüzen bir boncuk bulun ve deklanşörü açacak ve yakalama lazerinin numuneye çarpmasına izin verecek olan Tuzak Deklanşörü düğmesine tıklayarak yakalayın. Ekrandaki Tuzak imlecine tıklayın ve yakalama lazerinin konumunu hareket ettirmek için sürükleyin. Sıkıştıktan sonra, voltaj ölçümlerini kuvvet ve yer değiştirmeyle ilişkilendirmek için boncuğu kalibre edin.
- Kalibrasyon düğmesine tıklayın. Güç spektrumu analizine dayalı kalibrasyon rutinini ayarlayın ve X, Y ve Z yönleri için yazılımdaki köşe frekansını sığdırın (Ek Şekil S6).
- Ayarlar'a tıklayın. Boncuk çapını (1.000 nm) yazın ve yazılım penceresinin sol alt köşesinde bulunan aşamanın sıcaklığını yazın. (bkz. Ek Şekil S6).
- Tuzak 1'e tıklayın. X Sinyali'ne tıklayın. Köşe frekansına sığdırmak için Çalıştır'a tıklayın. İşlev uyumunu optimize etmek için pencerenin içine tıklayıp sürükleyin. Hassasiyet ve sertlik değerleri için Kullan'a tıklayın. Değerleri Kabul Et'e tıklayın. Y ve Z sinyalleri için bu işlemi tekrarlayın. Pencereyi kapatın. (bkz. Ek Şekil S6).
NOT: Diğer optik yakalama sistemlerinde veya kullanıcı tarafından sağlam bir şekilde test edilmiş özel yapım sistemlerde, örneğin eşit bölümleme yöntemi veya sürükleme kuvveti yöntemi gibi boncuk kalibrasyon rutinleri de kabul edilebilir57,58'dir. - Kapak kapağının yüzeyindeki AF'lere bağlı boncukları arayarak bir aktomiyozin demeti bulun.
- Diğer yüzen boncuklar tarafından kalabalık olmayan bir boncuk tespit edildiğinde, bir demetin varlığını doğrulamak için floresan görüntüleme ile etrafındaki AF'leri gözlemleyin.
- Her iki floresan AF'yi de birlikte lokalize ederek bir demetin mevcut olduğunu doğrulayın. Beyaz ışık kaynağını açın ve tareti çevirerek her bir aktin filamentini görüntülemek için uygun filtre küpünü kullanın (Alexa Fluor 488 ve rodamin uyarımı için sırasıyla 488 nm ve 532 nm uyarma filtresi küpleri). Bkz. Ek Şekil S4.
NOT: Tek AF'lerin floresan yoğunluğunu doğrulamak için yapılan bir kontrol deneyi, tek bir 488 ve tek rodamin etiketli filamentlerden oluşan veya kullanıcının kullanmayı seçtiği florofor kümesine uygulanabilir demetleri tanımlamada yararlı olabilir. - Doğrulandıktan sonra, Deklanşör Kapanı düğmesine tıklayarak paketin üst filamentine bağlı boncuğu yakalayın.
- Osiloskop düğmesine tıklayarak verileri kaydetmek için ekrandaki kontrolleri kullanın (Ek Şekil S7). Verileri kaydetmeden ölçümleri görselleştirmek için Başlat'a tıklayın. Tüm verileri kaydetmek için Otomatik Kaydet'e tıklayın. Ölçümleri kaydetmek için Kaydı Başlat'a tıklayın. X sinyali veya Y sinyali açılır menüsünden seçim yaparak hangi verilerin gerçek zamanlı olarak (konum, kuvvet, x yönü, y yönü) görselleştirileceğini seçin. xdirection'ın soldan sağa ve y yönünün ekranda yukarı ve aşağı olduğunu unutmayın. Bkz. Ek Şekil S7.
NOT: Veriler .out dosyaları olarak kaydedilir ve her yön için zaman, voltaj, yer değiştirme ve kuvvet içerir. Bu dosyalar görselleştirme ve analiz için diğer yazılımlara aktarılabilir.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Aktomiyozin demet sistemlerini içeren akış hücreleri, mikroskop sürgüsü ve çift taraflı yapışkan banttan yapılmış bir kanalla ayrılmış kazınmış bir kapak kaymasından oluşan standart bir tasarıma sahiptir (Şekil 1). Tahlil daha sonra protokolde açıklandığı gibi aşamalı tanıtımlar kullanılarak kapaktan yukarı doğru oluşturulur. Son tahlil, şablon rodamin etiketli aktin filamentlerinden oluşur; istenen miyozin konsantrasyonu ( Şekil 2 ve Şekil 3'teki temsili sonuçlar için 1 μM kullanılmıştır); biyotinillenmiş, Alexa Fluor 488 etiketli aktin filamentleri; 1 μm streptavidin boncuklar; oksijen süpürme sistemi; ATP; ve APB arabelleği. Akış hücresi başına çoklu demetler oluşturulacak ve yukarıda açıklanan aktin konsantrasyonları, istenmeyen etkileşimlerin olmamasını sağlamak için demetler arasında yeterli boşluk bırakacaktır. Bu aynı zamanda veri toplama verimliliğini artırmak için akış hücresi başına çoklu kuvvet ölçümleri elde etmeyi kolaylaştırır. Kuvvet profilleri bir akış hücresi içinde ve akış hücresinden akış hücresine tekrarlanabilir olmalıdır.
Yukarıdaki protokol ticari bir optik yakalama kurulumunun kullanımına yönelik olsa da, burada sunulan akış hücresi ve tahlil, mikroskop veya mikroskop aşaması ile birleştirilmiş ve floresan görüntüleme yeteneklerine sahip farklı bir ticari cihaz veya özel yapım optik yakalama kurulumu için kolayca kullanılabilir. Yukarıdaki protokole göre tüm akış hücresi ilaveleri tamamlandıktan sonra, slayttaki aktomiyozin demetleri (Şekil 1) hemen ölçüm için hazırdır. Akış hücresi optik tuzak mikroskobu aşamasına eklenir, çoklu boncuk kalibrasyon ölçümleri elde edilir ve demet filamentlerinin floresan kolokalizasyonu yoluyla demetler tanımlanır. Bir demete bağlı bir boncuk sıkışır ve yer değiştirme ve karşılık gelen kuvvet ölçümü başlar. Kullanıcı, bilgisayar monitöründe gerçek zamanlı olarak veri alımını gözlemleyebilir. Akış hücresinde kullanılan miyozin konsantrasyonuna bağlı olarak, demet hemen önemli bir hareket sergilemeye başlayabilir veya yer değiştirme / kuvvette etkili bir artış görmek için 30 s-1 dakika sürebilir.
Şekil 3A'da, miyozin motorlarının kuvvette sabit bir rampa sergilediği ve ardından bir plato izlediği temsili bir kuvvet izi gösterilmiştir. Bu tür izlerin 2-5 dakika içinde geliştiğini görmek tipiktir. Bununla birlikte, herhangi bir net kuvvet üretmeyen aktomiyozin demetlerini ölçmek de mümkündür (Şekil 3B). Bu izler temel gürültü olarak görünür veya 90'ların üzerinde kuvvette önemli bir net artış göstermez. Bu muhtemelen, üretken kaymaya izin vermeyen düşük bir yerel motor konsantrasyonundan veya demet, filamentlerin artı ve eksi uçlarının hizalandığı olumsuz bir paralel yönelimde olmasından kaynaklanmaktadır.
Akış hücresinin içeriği, olay aydınlatması ve yakalama lazerinden kaynaklanan bozulmaya, zaman içinde slaytta yerel ısınmaya ve radikal oksijen türlerinin üretilmesine duyarlı olabileceğinden, aynı akış hücresinin 1 saatten fazla kullanılmaması şiddetle tavsiye edilir. Maksimum verimlilik için, veri elde ederken başka bir tahlilin kuluçkaya yatırılması önerilir. Yer değiştirme/kuvvet izleme, optik yakalama yazılımından Excel, Matlab, Igor veya diğer veri yönetimi programlarına daha fazla filtreleme ve analiz için aktarılabilir. Bu tür optik yakalama topluluğu / demet deneylerinden çıkarılabilecek veriler, değişen tahlil koşulları altında farklı tipte kuvvet oluşturma profillerini (taban çizgisi, rampa / plato), kuvvet üretim hızı, maksimum kuvvet üretimi, adım boyutları ve adımlar veya adım ekipleri arasındaki bekleme süreleri boyunca kinetik ve adım atma davranışını ve görev oranını içerir. Kullanıcı ayrıca, farklı tipte miyozin motorlarının eklenmesinin, aktin bağlayıcı proteinlerin eklenmesinin veya tampon koşullarının değiştirilmesinin bu topluluk kuvveti oluşturma özelliklerini nasıl etkilediğini karşılaştırmak için tahlil koşullarını değiştirebilir.
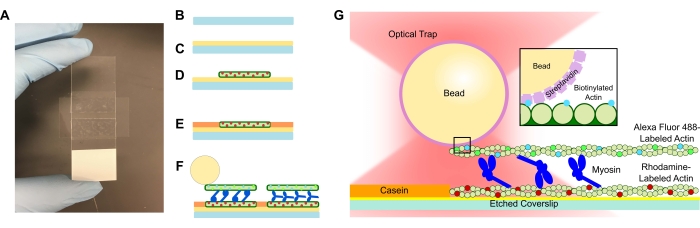
Resim 1: Tahlil şeması. (A-C) Kazınmış örtüler poli-L-lizin ile kaplanır ve çift taraflı bant ve mikroskop sürgüsü kullanılarak akış hücresini oluşturmak için kullanılır. Protokolde açıklanan zamanlanmış tanıtımlar ve inkübasyon adımları, şablon veya alt filament (D) olarak rodamin etiketli falloidin stabilize edilmiş aktin, ardından spesifik olmayan bağlanmayı önlemek için kazein blokajı (E) ve (F) Alexa Fluor 488 faloidin stabilize edilmiş biyotinillenmiş aktin kargo veya üst filament olarak ve filamentleri birbirinden ayıran ve ATP tanıtıldığında kuvvet üreten miyozin II ekipleri ile sonuçlanır. Motorların geometrisi ve demet içindeki çapraz bağlamanın doğası, tuz konsantrasyonu59 gibi farklı koşullar altında değişebilir. Önceki çalışmalar, miyozin kuyruğu alanının aktin filamentleri ve yavaş topluluk hareketliliği ile etkileşime girme yeteneğine sahip olduğunu göstermiştir46. Bununla birlikte, ağır meromiyozin deneylerinde miyozin kafaları, her bir başın bitişik aktin filamentlerine bağlandığını göstermektedir60. (G) Streptavidin boncukları, tuzak için optik tutamak olarak kullanılır ve yalnızca kargo biyotinile aktin filamentine bağlanır, bu da slaytta uygun demetlerin oluştuğunu doğrulamaya yardımcı olur. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.

Şekil 2: Floresan aktomiyozin demetleri. Aktin filamentlerinin ve demetlerinin Şekil 1'de sunulan demet tahlili içindeki dört farklı karşılaşması. Alexa Fluor 488 falloidin kanallı üst kargo biyotinile aktin filamenti solda, alt şablon ise rodamin falloidin kanallı aktin filamenti sağda gösterilmiştir. Alt kısımda, aynı şekil gözü yönlendirmeye yardımcı olmak için üst üste bindirilmiş renkli çizgilerle gösterilmiştir. (A) Bir üst aktin filament, bir alt aktin filamentinin yakınında bulunur, ancak eksik bir kaplamaya sahiptir. Bu, paket denemeleri için kullanılmaz. (B) Üst ve alt aktin filamentleri birlikte lokalize edilir ve her bir filamentin yoğunluğu, bunların demet içindeki her bir filament olduğunu doğrular. Bu, paket deneyleri için iyi bir aday olacaktır. (C) Altta kendinden montajlı rodamin filamentlerinden oluşan büyük bir demet bulunur. Kolokalize edilmiş karşılık gelen bir üst aktin filamenti olsa da, çok fazla alt filament mevcuttur; bu nedenle, paket deneyleri için kullanılmaz. Bu aynı zamanda, aynı tipteki çoklu aktin filamentleri paketlendiğinde, floresan yoğunluğunun nasıl arttığının bir örneğidir. Kullanıcı bunu, aynı filament tipindeki demetlere karşı tek filamentleri yargılamak için bir gösterge olarak kullanabilir. (D) Karşılık gelen üst filament içermeyen bir alt filament bulunur ve ayrıca kanama olmadığını doğrular. Bu, paket denemeleri için kullanılmaz. Alexa Fluor 488 kanalındaki filamentlerin yoğunluğunun düşük olduğunu ve bunun kullanılan filtre setinden kaynaklandığına inanıyoruz (Zeiss'tan Filtre Seti 09). Rodamin kanalı için kullanılan filtre seti, Zeiss'ın Filtre Seti 43'tür. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.

Resim 3: Myosin II topluluk kuvveti üretimi. İnşa edilmiş in vitro aktin yapısal hiyerarşi içinde kuvvet üreten iskelet miyozin II motorlarının temsili izleri. Miyozin motorları, bir platoya ulaşılana ve kuvvet sürdürülene (A) veya taban çizgisine (B) yakın antagonizma deneyimleyene kadar kolektif ve üretken bir şekilde kuvvet üretmek için birlikte çalışırlar. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
Ek Şekil S1: Bruker / JPK Nanotracker2 optik tuzağı. (A) Bilgisayar monitörü. (B) Bilgisayar klavyesi. (C) Bilgisayar kulesi. (D) Denetleyici kutusu. (E) Lazer güç kaynağı. (F) Optik tuzak optik kutusu. (G) Ters mikroskop. (H) Mikroskop aşamasına kapı. (I) Parlak alan ve diferansiyel girişimli kontrast görüntüleme arasında geçiş yapmak için polarizör kaydırıcısı. Bu Dosyayı indirmek için lütfen tıklayınız.
Ek Şekil S2: Optik tuzak için uzaktan kumanda. (A) Motorlu sahneyi konumlandırmak için tuş takımı. (B-C) Tuzak konumunu ayarlayın. (D) A, X ve B sırasıyla ana deklanşörü açıp kapatır, 1 deklanşörü yakalar ve 2 deklanşörü yakalar. (E) Logitech düğmesi, oyun kumandasını uyandırmak için kullanılır. (F) Tuzak hedefini konumlandırmak için kullanılan yukarı ve aşağı düğmeleri. (G) Algılama hedefini konumlandırmak için kullanılan yukarı ve aşağı düğmeleri. Uzaktan kumandanın gerekli olmadığını ve tüm bu manipülasyonların yazılımda gerçekleştirilebileceğini unutmayın. Bununla birlikte, mikroskop sahne ortamına bakarken hedefleri ve sahne konumunu kontrol edebilmek uygundur. Bu Dosyayı indirmek için lütfen tıklayınız.
Ek Şekil S3: Optik tuzak için floresan modülü. 89North FotoFlor floresan beyaz ışık kaynağı, ters çevrilmiş mikroskobun arkasına bağlanır. Bir geçiş düğmesiyle (ok) açılır ve kapatılır. Bu Dosyayı indirmek için lütfen tıklayınız.
Ek Şekil S4: Floresan filtre küp tareti. Taret (ok), DIC, rodamin veya Alexa Fluor 488 boyalarında görüntüleme için gerekli filtre küpünü kullanacak şekilde döndürülebilir. Farklı floroforlar kullanmak üzere kurulumu özelleştirmek için filtre küplerinin kapatılabileceğini unutmayın. Bu Dosyayı indirmek için lütfen tıklayınız.
Ek Şekil S5: Nanotracker2 yazılımı. (A) Lazer güç düğmesi ve kontrolü. (B) Objektif konumlandırma penceresi. Yön okları, algılama (üstte) ve yakalama (altta) hedeflerini hareket ettirmek için kullanılır. Çift oklar hedefleri daha yüksek bir hızda hareket ettirir. Sol alttaki mavi ve kırmızı düğme hedefleri birbirinden ayırır ve orijinal konumlarına geri çeker. Bu, mikroskop aşamasının içine ve dışına numune alırken gereklidir. Soldan gelen üçüncü düğme, hedefler ve asma kilit simgesi ile hedefleri "eşleştirir", böylece hem odakta olduklarında hem de Kohler aydınlatmasına ulaştıklarında, kullanıcı hem yakalama hem de algılama hedeflerini z ekseninde yukarı ve aşağı hareket ettirebilir. (C) Mikroskop aşamasını x ve y ekseninde hareket ettirmek için kullanılan numune konumlandırma penceresi. Çift oklar sahne alanını daha yüksek bir hızda hareket ettirir. Bu pencere, üst menüdeki yukarı/aşağı ve sol/sağ ok simgesine tıklanarak etkinleştirilir. (D) Kamera görselleştirme penceresi. Anahtar simgesi, özelleştirilmiş görüntüleme koşullarını ayarlamak için kullanılabilir. Bu pencere, üst menüdeki Kamera simgesine tıklanarak etkinleştirilir. (E) Mikroskop aydınlatma penceresi. Bu pencere, üst menüdeki Ampul simgesine tıklayarak etkinleştirilir. Bu Dosyayı indirmek için lütfen tıklayınız.
Ek Şekil S6: Kalibrasyon penceresi. (A) Bu pencere boncuk kalibrasyonu için kullanılır ve üst menüdeki Cal simgesine tıklanarak etkinleştirilir. Bir boncuğu kalibre etmek için, köşe frekansının en iyi uyumu x, y ve z sinyallerinde gerçekleştirilir. (B) Her sinyal için sol üstteki uygun sinyal düğmesini seçin. (C) Çalıştır'a tıklayın ve yeşil pencerede (D) tıklayıp sürükleyerek uyumu optimize edin. (E) Uyumdan memnun kaldıktan sonra, hassasiyet ve sertlik için Kullan'a tıklayın. Bu, nanometrelerde yer değiştirmenin ve pikonewtonlarda kuvvetin kaydedilmesine izin verecektir. (F) Ardından, sol alttaki Değerleri Kabul Et'e tıklayın. Y ve z yönleri için tekrarlayın. Bu Dosyayı indirmek için lütfen tıklayınız.
Ek Şekil S7: Veri Toplama penceresi. Bu pencere, konum ve kuvvet verilerini almak için kullanılır ve kullanıcının ölçümleri gerçek zamanlı olarak görmesini sağlar. (A) Bu pencere, üst menüdeki x,t simgesine tıklanarak etkinleştirilir. (B) Kullanıcı x ve y sinyallerini görüntüleme arasında geçiş yapabilir. (C) Verileri görselleştirmeye başlamak için Başlat'a tıklayın. Verileri kaydetmek için Otomatik Kaydet'e tıklayın. Verileri kaydetmeye ve kaydetmeye başlamak için Kaydı Başlat'a tıklayın. Bu Dosyayı indirmek için lütfen tıklayınız.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Aktin filamentlerle etkileşime giren miyozin topluluklarının dinamiklerini araştırmak için floresan görüntüleme ile birlikte optik cımbız kullanan in vitro bir çalışma yapıldı. Aktin-miyosin-aktin demetleri, kas miyozin II, demetin dibinde ve kapak kayma yüzeyinde rodamin aktin ve demetin üstünde 488 etiketli, biyotinile aktin filamentleri kullanılarak birleştirildi. Tavşan kasından elde edilen aktin proteini, genel aktin tamponları (GAB) ve aktin polimerize edici tamponlar (APB) kullanılarak polimerize edildi ve stabilize edildi. GAB ve APB, laboratuvarda ATP, FC tamponu ve TC tamponu kullanılarak her gün taze olarak hazırlanmalıdır. Aktin-miyosin-aktin sandviçlerini oluşturmak için kas miyozin II kullanıldı. Faloidin, aktin filamentlerinin floresan boyanması ve in vitro stabilizasyonu için kullanılmıştır.
Myosin aktivitesi, daha önce 46,47'de yayınlandığı gibi standart bir kayma filament testi yapılarak doğrulanabilir. Myosin II ve alt parçaları, örtü kayması yüzeyine çeşitli yönlerde bağlanabilir ve kuyruk alanının varlığı, ağır meromiyozin46,48,49 kullanan tahlillere kıyasla filament kaymasını yavaşlatabilir. Bununla birlikte, kayma ve yüzey hareketi hala gözlemlenebilir. Miyozin aktivitesinin daha belirgin bir gösterimi, daha uzun aktin filamentlerinin daha sonra çok yönlü olarak kayan daha küçük parçalara ayrıldığı yerlerde gözlenebilen aktif aktin filament kırılmasıdır. Bu, yüzeydeki aktif motorların yüksek konsantrasyonundan kaynaklanır, birden fazla laboratuvar tarafından gözlemlenmiştir ve aktif miyozin motorları 42,50,51,52,53,54 olmadan gerçekleşmez. Ayrıca, burada sunulan demet testi, öncelikle bir cam kapak kayması üzerindeki motor bağlama yönelimlerinin çeşitliliği gibi kayma filament testi ile ilişkili olan hareketlilik sorunlarının hafifletilmesine yardımcı olur, çünkü demet testi, cam yüzeyin kazein blokajını içerir, böylece motorlar 47,55,56 demet içinde bağlanır.
İlk adım, bir akış hücresindeki poli-L-lizin kaplı bir örtü kaymasına alt veya şablon filament olarak rodamin aktin filamentleri eklemektir. Poli-L-lizin, aktin bağlanmasını teşvik etmek için kullanılır, çünkü poli-lizin pozitif yüklüyken, aktin negatif yüklere sahiptir ve önceki sitoiskelet in vitro tahlil preparatlarında kullanılmıştır61,62,63. Demet oluşumundan önce, aktin konsantrasyonunu optimize etmek için bir akış hücresine farklı aktin seyreltmeleri eklenmiştir. Bu durumda, stoktan 600x, demet oluşumu için yeterli sayıda şablon filamenti veren, ancak demetlerin bireyselleştirilmesi için yeterli aralıkla optimum seyreltmeydi. Seyreltme APB tamponu kullanılarak gerçekleştirildi. Rodamin aktininin eklenmesini, yüzeyi bloke etmek ve spesifik olmayan bağlanmayı önlemek için bir kazein tabakası izledi. Akış hücresi 30 dakika boyunca inkübe edildi ve bağlanmamış aktin filamentlerini yıkamak için inkübasyondan sonra tamponla yıkandı. Son olarak, aktin-miyozin demeti oluşumunu kolaylaştırmak için akış hücresine miyosin, 488 / biyotin aktin ve streptavidin kaplı boncukların bir kombinasyonu eklendi. Boncuk konsantrasyonu, yüzeye bağlı demetleri bağlamak için yeterli ve kalibrasyonu kolaylaştırmak için süspansiyonda yeterli olacak şekilde olmalıdır. Bununla birlikte, bir boncuk konsantrasyonunun çok yüksek olması, lazer tuzağına düşen ve ölçümü bozan komşu boncuklar nedeniyle yakalama deneyleri sırasında zorluğa neden olabilir. Myosin motorları, slayta enjekte edilmeden hemen önce kombinasyona eklenir, böylece miyozin motorları kargo veya üst biyotinile aktin filamenti ile önleyici olarak toplanmaz ve böylece alt rodamini biyotinillenmiş aktin filamentlerini demetlemek için bağlar.
NT2 optik yakalama sistemi, kombine parlak alan, diferansiyel girişim kontrastı (DIC) ve epifloresan görüntüleme yöntemlerine sahip ticari bir optik tuzaktır. 100x/NA 1.46 ve 63x/NA 1.0 suya daldırma yakalama ve algılama hedeflerine sahip bir Zeiss AxioObserver 3 ters mikroskop ile birleştirilmiştir. Sistem, bir lazer tuzağının tıklama ve sürükleme yakalama özelliği ile donatılmıştır ve daha önce listelenen yöntemlerden herhangi birinde görüntüleme yaparken kullanılabilir. Oluşan demetler floresan görüntüleme kullanılarak tespit edilir ve doğrulanır. Uygun filtre küplerine (GFP/FITC ve TRITC/CY3) sahip beyaz bir ışık kaynağına sahip olmak, filament görüntüleme arasında hızlı geçiş yapılmasını sağlar. Kolokalize AF'ler, optik cımbız kullanarak her kuvvet ölçümünü yapmadan önce AF'leri farklı uyarma dalga boylarında görselleştirerek doğrulandı. Filamentler, bir oksijen süpürme reaktifi ile bile hızlı bir şekilde fotobeyazlatma yapabildiğinden, araştırmacıların paket deneylerini gerçekleştirmeden önce yoğunluk ve maruz kalma süresi gibi görselleştirme parametrelerini optimize etmeleri önerilmektedir.
Biyotinile kargo aktin filamentini bağlamak ve bir kuvvet dönüştürücü olarak miyozin kuvvet üretimini aktive etmek için ATP varlığında streptavidin boncuklarını kullanarak kuvvet ölçümlerini almak için optik yakalama kullanılmıştır. Optik yakalama ile elde edilen yer değiştirme ve kuvvete karşı zaman verileri, analiz için yakalama yazılımından çıkarılmıştır. Bununla birlikte, ticari yakalama yazılımı ayrıca kullanılabilecek analiz rutinleri sağlar veya diğer programlardaki özel algoritmalar, tuzak verilerini görselleştirmek ve analiz etmek için kullanıcı tarafından programlanabilir. Özel optik yakalama sistemlerinde, kullanıcı filtrelere sahip beyaz bir ışık kaynağı yerine uyarma lazerlerine sahip olabilir ve bunlar da kullanımı kabul edilebilir. Ayrıca, floresan boyalar, emisyon spektrumları üst üste binmemesi ve kanamaya neden olmaması durumunda kullanıcının sahip olabileceği mevcut ekipmana uyacak şekilde değiştirilebilir.
Sunulan tahlilin, aktomiyozin topluluğu mekaniği alanındaki araştırma sorusuna bağlı olarak kullanıcı tarafından daha da özelleştirilebilen bir temel tahlil olduğunu not ediyoruz. Genel iş akışı, mitotik iğ 32,61,63,64,65,66'nın minimal modellerini oluşturan mikrotübül demet tahlilleri gibi ilgi çekici olabilecek diğer in vitro sitoiskelet topluluğu sistemlerine de uygulanabilir. Değişiklikler, kullanıcının mevcut kurulumuna uygun florofor etiketlerini değiştirmeyi içerebilir, ancak bunlarla sınırlı değildir; miyozin konsantrasyonunu, yapısını veya izotipini değiştirmek; ve diğer yönlerin yanı sıra titre tampon koşulları.
Bu tahlil yapılırken potansiyel zorluklar mümkündür. Aktin-miyozin demetlerini oluştururken, aktin demetleri içindeki miyozin konsantrasyonu slayt boyunca homojen olmayabilir. Buna uyum sağlamak için, motor dağılımı ve kuvvet oluşturma profillerinin uygun şekilde örneklendiğinden emin olmak için tüm slayt boyunca birden fazla demet ölçülecektir. Kuvvet verilerinin yorumlanması için bunun gerekli olup olmadığını bilmek de zordur. Bu nedenle, her paket için birden fazla deneme yapılmalıdır. Aktin filament ucu etiketlemesi, optik yakalama sapından daha küçük boyutlu floresan jelsolin veya jelsolin kaplı boncuklar aracılığıyla da dahil edilebilir. Floresan görüntüleme, demet oryantasyonunu çıkarmak için x ve y bileşen kuvvetlerine bakmak için de kullanılabilir. Ayrıca, miyozin agregasyon durumu, KCl'nin hızlı seyreltilmesi üzerine meydana gelen kalın filamentlerin oluşumu ile tamponun iyonik gücünden oldukça etkilendiğinden, tampon tuzu konsantrasyonu uygun şekilde izlenmelidir67,68.
Kayma testleri gibi diğer in vitro yöntemleri kullanan önceki çalışmalar, miyozin alanlarının rolünü tanımlamak ve miyozin ile diğer aktin bağlayıcı proteinler arasındaki konfigürasyonu ve etkileşimleri incelemek için yardımcı olmuştur. Bununla birlikte, bu yöntemlerin, miyozinin sert bir yüzeye bağlanmasının, miyozin motorları arasındaki koordinasyon potansiyelini sınırlayacağı ve böylece motor topluluğunun yüksek veya düşük bir görev oranı modunda olup olmadığını belirlemek için meydana gelen mekanosensing geri bildirimini sınırlayacağı için bir dezavantajı vardır. yüksek veya düşük görev oranı modu 33,35,41,69. Ayrıca, tek miyozin motor ağları ile optik yakalama, miyozin motorlarının birbirleriyle ve aktin filamentleriyle nasıl etkileşime girdiğine dair net bir anlayış sağlamaz. Burada geliştirilen protokol, miyozin motor topluluk dinamiklerinin uyumlu, hiyerarşik bir aktin ağı içinde araştırılmasına izin verir. Ayrıca, sistematik araştırmaya izin vermek için diğer yönlerin yanı sıra konsantrasyon, izoform ve tampon ortamı gibi motor-filament topluluk özellikleri açısından da özelleştirilebilir. Sunulan protokol, daha karmaşık aktomiyozin ağlarının gelecekteki çalışmaları için bir platformdur ve geleneksel olarak tek moleküllü çalışmalar için kullanılan optik yakalama ile kolaylaştırılan yer değiştirme ve kuvvet oluşturma ölçümlerinin hassasiyetini korur.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Yazarların beyan edecekleri herhangi bir çıkar çatışması yoktur.
Acknowledgments
Bu çalışma kısmen Mississippi Üniversitesi Lisansüstü Öğrenci Konseyi Araştırma Bursu (OA), Mississippi Üniversitesi Sally McDonnell-Barksdale Honors College (JCW, JER), NNX15AH78H (JCW, DNR) hibe numarası altında Mississippi Uzay Hibe Konsorsiyumu ve 848586 hibe numarası (DNR) altında Amerikan Kalp Derneği tarafından desteklenmektedir.
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
