

Summary
היווצרות של צרורות actomyosin במבחנה ומדידת ייצור כוח אנסמבל מיוזין באמצעות פינצטה אופטית מוצג ונדון.
Abstract
מיוסינים הם חלבונים מוטוריים המבצעים הידרוליזה של ATP כדי לצעוד לאורך מסלולי נימה אקטין (AF) והם חיוניים בתהליכים תאיים כגון תנועתיות והתכווצות שרירים. כדי להבין את מנגנוני ייצור הכוח שלהם, מיוזין II נחקר הן ברמת המולקולה הבודדת (SM) והן כצוותים של מנועים במבחנה באמצעות שיטות ביופיזיות כגון לכידה אופטית.
מחקרים אלה הראו שהתנהגות של יצירת כוח מיוזין יכולה להיות שונה מאוד כאשר עוברים מרמת המולקולה הבודדת בסידור של שלושה חרוזים לקבוצות של מנועים העובדים יחד על משטח חרוז קשיח או כיסוי בסידור גלישה. עם זאת, מבני בדיקה אלה אינם מאפשרים להעריך את הדינמיקה הקבוצתית של מיוזין בתוך היררכיה מבנית ויסקו-אלסטית כפי שהיו עושים בתוך תא. פיתחנו שיטה המשתמשת בפינצטה אופטית כדי לחקור את המכניקה של יצירת כוח על ידי הרכבי מיוזין המקיימים אינטראקציה עם חוטי אקטין מרובים.
צרורות אקטומיוזין אלה מאפשרים חקירה בסביבה היררכית ותואמת, הלוכדת תקשורת מוטורית ותפוקת כוח אנסמבל. האופי הניתן להתאמה אישית של הבדיקה מאפשר לשנות את תנאי הניסוי כדי להבין כיצד שינויים בהרכב המיוזין, בצרור חוטי האקטין או בסביבה הסובבת גורמים לתפוקות כוח שונות.
Introduction
חלבונים מוטוריים חיוניים לחיים, וממירים אנרגיה כימית לעבודה מכנית 1,2,3. מנועי מיוזין מתקשרים עם חוטי אקטין על ידי נקיטת צעדים לאורך החוטים הדומים למסלול, והדינמיקה של רשתות אקטין-מיוזין מבצעת התכווצות שרירים, תנועתיות תאים, טבעת התכווצות במהלך ציטוקינזיס ותנועת מטען בתוך התא, בין שאר המשימות החיוניות 3,4,5,6,7,8 . מאחר שלמיוסינים יש כל כך הרבה תפקידים חיוניים, כשל בתפקוד של רשת המיוזין-אקטין יכול להוביל להתפתחות מחלות, כגון מוטציות בשרשרת הכבדה של המיוזין הגורמות להיפר-התכווצויות יתר של הלב בקרדיומיופתיה היפרטרופית (HCM)9,10,11,12,13,14 . בהתכווצות שרירים, מנועי מיוזין בודדים משתפים פעולה זה עם זה על ידי עבודה כאנסמבל כדי לספק את האנרגיה המכנית הנדרשת המבצעת את ההחלקה היחסית של AFs 4,15,16,17,18. מנועי מיוזין יוצרים גשרים צולבים בין AFs ומשתמשים בשינויים קונפורמיים עקב המחזור המכנוכימי שלהם כדי לנוע באופן קולקטיבי לעבר הקצה התיל של החוטים המיושרים 17,18,19,20,21.
פיתוח מבחני תנועתיות כמותיים במבחנה ברמת ה-SM תוך שימוש בטכניקות כגון השמנה אופטית הקל על איסוף פרטים חסרי תקדים על האופן שבו מנועי מיוזין בודדים מתפקדים, כולל מדידת ייצור כוח SM וגדלי צעדים 22,23,24,25,26,27,28,29,30 . Finer et al. פיתחו את בדיקת הלכידה האופטית "שלושה חרוזים" או "משקולת" כדי לחקור את מכניקת ייצור הכוח של מנועי מיוזין II בודדים23,31. מכיוון שמיוזין שריר II עובד בצוותים כדי לכווץ AFs אך אינו מעובד ברמת SM, היה צורך לארגן מחדש את כיוון בדיקת הלכידה האופטית מהגישה הקלאסית של חרוזים הקשורים למנוע32. כדי ליצור את בדיקת המשקולת, שתי מלכודות אופטיות שימשו להחזקת AF מעל מנוע מיוזין הקשור לחרוז המחובר לכיסוי, ותפוקת הכוח על ידי המנוע היחיד נמדדה באמצעות תנועות של המיקוד האוטומטי בתוך מלכודת23.
עם זאת, כוחות SM ושימוש בכיוון בדיקה של מנוע יחיד/נימה בודדת אינם נותנים תמונה מלאה על יצירת כוח ברמת המערכת מכיוון שחלבונים מוטוריים רבים, כולל מיוזין II, אינם פועלים בבידוד ולעתים קרובות אינם מתפקדים כסכום חלקיהם 15,16,17,32,33,34,35,36 . מבנים מורכבים יותר הכוללים יותר ממנוע אחד המקיים אינטראקציה עם יותר מחוט אחד נחוצים כדי להבין טוב יותר את הסינרגיה של רשתות המיוזין וחוטי האקטין15,32. כיוון בדיקת המשקולת נוצל כדי לחקור יצירת כוח אנסמבל קטן על ידי חיבור מיוזינים מרובים לחרוז או שימוש בחוט עבה מיוזין המחובר למשטח ומאפשר למנועים לקיים אינטראקציה עם המיקוד האוטומטיהתלוי 4,23,34,37,38,39,40.
מבחני אנסמבל קטנים אחרים כוללים בדיקת גלישה של נימה במבחנה שבה מנועי מיוזין מצופים על משטח כיסוי, וחרוז הקשור ל- AF משמש כדי לחקור את הכוח שנוצר על ידי צוות המנועים 4,35,36,38,39,40,41,42,43 . בשני המקרים הללו, המיוזינים קשורים למשטח קשיח - חרוז או כיסוי - ומשתמשים במיקוד אוטומטי אחד. במקרים אלה, המנועים אינם מסוגלים לנוע בחופשיות או לתקשר זה עם זה, וגם קיום מיוסינים הקשורים באופן נוקשה אינו משקף את הסביבה התואמת, ההיררכית, שבה המנועים יעבדו יחד בסרקומר32. מחקרים קודמים הציעו כי מיוזין II יכול לחוש את סביבתו ולהסתגל בהתאם לשינויים בתנאי ריכוז ויסקואלסטיים או מוטוריים על ידי שינוי מאפיינים כגון ייצור כוח ויחס חובה41,44,45. לפיכך, יש צורך לפתח מבחן לכידה אופטי המטפח ולוכד תקשורת מוטורית ותאימות מערכת כדי לצייר תמונה מציאותית יותר של היסודות המכניסטיים של יצירת כוח הרכב מיוזין II.
כאן, פיתחנו שיטה לשילוב מבנה היררכי במבחנה עם לכידה אופטית על ידי יצירת צרורות אקטומיוזין או סנדוויצ'ים המורכבים ממנועי מיוזין מרובים המקיימים אינטראקציה בין שני חוטי אקטין. לגיאומטריה מודולרית זו יש את היכולת לחקור ישירות כיצד גורמים מולקולריים וסביבתיים משפיעים על יצירת כוח מיוזין בהרכב. יתר על כן, לחקר מנגנוני יצירת הכוח באמצעות הרכבי אקטין-מיוזין אלה יש פוטנציאל לסייע במידול ובהבנת האופן שבו משימות תאיות בקנה מידה גדול, כגון התכווצות שרירים, מתפשטות מהרמה המולקולרית 9,10,13.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. כיסויי תחריט
- יש להמיס 100 גרם של KOH ב-300 מ"ל של 100% אתנול בכוס של 1,000 מ"ל. מערבבים עם מוט ערבוב עד שרוב ה-KOH מתמוסס.
אזהרה: תמיסת KOH מרוכזת עלולה לגרום לכוויות ולנזק לבגדים. יש ללבוש כפפות, להגן על העיניים ומעיל מעבדה. - הניחו את הכיסויים בנפרד במדפים לניקוי כיסויים.
הערה: המדפים מתוכננים עם חריצים המחזיקים כיסויים בודדים במרווחים זה מזה כדי לאפשר תחריט ושטיפה בכל פאה של הכיסוי, חורי ניקוז בתחתית ועשויים מחומר שיכול לעמוד בתנאי התחריט הקשים. הם יכולים להיות בהתאמה אישית או לרכוש באופן מסחרי. - הכינו ותייגו שלוש כוסות בגודל 1,000 מ"ל: אחת עם 300 מ"ל אתנול ושתי כוסות עם 300 מ"ל מים באוסמוזה הפוכה (RO).
הערה: כאן, מי RO מקורם במטהר מים במעבדה, אך ניתן גם לרכוש אותם באופן מסחרי אם מטהר מקומי אינו זמין. - מניחים כל אחד מארבעת הכוסות בסוניק אמבטיה כדי להתנתק במשך 5 דקות.
- לטבול מתלה של כיסויים בכוס של KOH ואתנול ו sonicate במשך 5 דקות.
- העבירו את מתלה הכיסויים מכוס KOH/אתנול לכוס האתנול בלבד. טובלים מעלה ומטה בכוס עד שאין חרוזים.
הערה: הקפידו לא להפריע לכיסויים או להפיל בכוח את המדף לתוך הכוס. פעולה זו תגרום לכיסויים לצאת מהמדף או לגרום להתזה כימית. - מעבירים בזהירות את מתלה הכיסויים מכוס האתנול לכוס מים, טובלים מעלה ומטה עד שאין חרוזים.
- הטביעו את מתלה הכיסויים בכוס המים שעדיין לא נעשה בה שימוש וסוניק שוב למשך 5 דקות.
- השתמשו בבקבוק כדי לרסס את מדף הכיסויים במים עד שהוא יורד מהכיסויים בצורה חלקה. חוזרים על הפעולה עם האתנול.
- מניחים את המדפים לייבוש בתנור בטמפרטורה של 90 מעלות צלזיוס למשך 20 דקות. אחסנו את מדפי הכיסויים החרוטים בטמפרטורת החדר במיכלים סגורים כדי למנוע זיהום לפני השימוש.
2. פילמור נימה אקטין
- הפוך את פתרון T
- בצינור חרוטי של 50 מ"ל, מוסיפים 3.94 גרם של Tris-HCl ו-0.147 גרם של CaCl2. מוסיפים מי RO כדי ליצור נפח כולל של 50 מ"ל ומערבבים היטב.
הערה: הריכוזים הסופיים של תמיסה T הם 500 mM Tris-HCl ו- 20 mM CaCl2, בהתאמה. - תייג את הצינור פתרון T ואחסן אותו ב 4 °C.
- בצינור חרוטי של 50 מ"ל, מוסיפים 3.94 גרם של Tris-HCl ו-0.147 גרם של CaCl2. מוסיפים מי RO כדי ליצור נפח כולל של 50 מ"ל ומערבבים היטב.
- צור מאגר TC
- מערבבים 40 מ"ל של מי RO ו-1.5 מ"ל של תמיסה T בצינור חרוטי של 50 מ"ל. שנה את ה- pH ל- 8.0 על-ידי הוספת כמויות קטנות של KOH מרוכז. מוסיפים מים כדי להפוך 50 מ"ל של הפתרון, ולאמת את ה- pH. התאם את ה- pH במידת הצורך.
הערה: מאגר TC הסופי מכיל 5 mM Tris-HCl ו- 0.2 mM CaCl2 ב- pH 8. - תייג את הצינור TC ואחסן אותו ב 4 °C.
- מערבבים 40 מ"ל של מי RO ו-1.5 מ"ל של תמיסה T בצינור חרוטי של 50 מ"ל. שנה את ה- pH ל- 8.0 על-ידי הוספת כמויות קטנות של KOH מרוכז. מוסיפים מים כדי להפוך 50 מ"ל של הפתרון, ולאמת את ה- pH. התאם את ה- pH במידת הצורך.
- צור מאגר FC
- הוסיפו 85 מ"ל של מי RO, 10 מ"ל של תמיסה T, 3.73 גרם של KCl ו-0.041 גרם של MgCl2 לבקבוק חיץ של 100 מ"ל. שנה את ה- pH ל- 7.5 על-ידי הוספת נפחים קטנים של KOH מרוכז. מוסיפים מים כדי ליצור נפח סופי של 100 מ"ל ולאמת את ה- pH.
הערה: מאגר FC הסופי מכיל 500 mM Tris-HCl, 500 mM KCl, 2 mM MgCl 2 ו- 2 mM CaCl2 ב- pH 7.5. - תייג את הצינור FC ואחסן אותו ב 4 °C.
- הוסיפו 85 מ"ל של מי RO, 10 מ"ל של תמיסה T, 3.73 גרם של KCl ו-0.041 גרם של MgCl2 לבקבוק חיץ של 100 מ"ל. שנה את ה- pH ל- 7.5 על-ידי הוספת נפחים קטנים של KOH מרוכז. מוסיפים מים כדי ליצור נפח סופי של 100 מ"ל ולאמת את ה- pH.
- הכן מאגר אקטין כללי (GAB).
- ערבבו 485 μL של מאגר TC, 10 μL של 10 mM ATP ו-5 μL של 50 mM DTT בצינור מיקרוצנטריפוגה.
הערה: תנאי החיץ הסופיים הם 5 mM Tris-HCl, 0.2 mM CaCl 2, 0.5 mM DTT ו-0.2 mM ATP. - תייג אותו כ- GAB ואחסן אותו ב- 4 °C.
- ערבבו 485 μL של מאגר TC, 10 μL של 10 mM ATP ו-5 μL של 50 mM DTT בצינור מיקרוצנטריפוגה.
- הכן את מאגר הפולימריזציה של אקטין (APB).
- ערבבו 455 μL של חיץ FC, 25 μL של 100 mM ATP, ו- 20 μL של 50 mM DTT בצינור מיקרוצנטריפוגה.
הערה: תנאי החיץ הסופיים הם 50 mM Tris-HCl, 500 mM KCl, 2 mM MgCl 2, 2 mM CaCl 22 mM DTT ו- 5 mM ATP. - תייג את הצינור כ-APB ואחסן אותו ב-4 מעלות צלזיוס.
- ערבבו 455 μL של חיץ FC, 25 μL של 100 mM ATP, ו- 20 μL של 50 mM DTT בצינור מיקרוצנטריפוגה.
- שחזור אקטין
- שחזרו את אקטין שרירי שלד הארנב על ידי הוספת 100 μL של מים שעברו דה-יוניזציה לבקבוקון של 1 מ"ג של אקטין ליופיליזציה. מערבבים היטב על ידי צנרת עדינה למעלה ולמטה. Aliquot לתוך דגימות 5 μL, להקפא, ולאחסן את 10 מ"ג / מ"ל אקטין aliquots ב -80 °C.
- שחזרו את אקטין שרירי שלד הארנב על ידי הוספת 20 מיקרון ליטר של מי RO. Aliquot לתוך דגימות 5 μL, הקפאה הצמדה, ולאחסן את 1 מ"ג / מ"ל ביוטינילציה אקטין aliquots ב -80 °C.
- פולימריזציה של אקטין ללא תווית עם ייצוב רודאמין פלואידין
- הפשירו בקבוקון אחד של 10 מ"ג/מ"ל אקטין ושמרו אותו על קרח.
- הכינו מאגר GAB טרי, הוסיפו 100 μL של GAB לאקטין אליקוט, וערבבו על ידי צנרת עדינה למעלה ולמטה. לדגור את התמיסה על הקרח במשך שעה אחת.
- הכינו APB טרי במהלך הדגירה. לאחר הדגירה, פילמר את האקטין לתוך חוטים על ידי הוספת 11 μL של APB לתמיסת אקטין. מערבבים היטב על ידי צנרת עדינה למעלה ולמטה. מניחים על הקרח למשך 20 דקות.
- הוסיפו 5 μL של פלואידין עם תווית רודמין לתמיסת חוטי אקטין פולימריים טריים. משאירים על הקרח בחושך למשך שעה.
- אחסנו את בקבוקון הרודמין אקטין עטוף ברדיד אלומיניום בחושך בטמפרטורה של 4 מעלות צלזיוס.
הערה: מומלץ להשתמש בחוטים אלה לתקופה מקסימלית של שבוע אחד. ניתן לאשר את איכות המיקוד האוטומטי בכל יום באמצעות הדמיה מהירה של תא זרימה המכיל רק הודעות מיקוד אוטומטי וצפייה בחוטים עקביים מדי יום.
- פולימריזציה של אקטין ביוטינילציה עם ייצוב אלפאלואידין אלקסה פלואור 488
- הפשירו בקבוקון אחד של 10 מ"ג/מ"ל אקטין ובקבוקון אחד של 1 מ"ג/מ"ל אקטין שעבר ביוטינילציה ושמרו אותם על קרח.
- הכינו מאגר GAB טרי.
- ערבבו את שני הבקבוקונים (שלב 2.8.1) ביחס אקטין 10:1 ביחס ביוטינילציה של אקטין. הוסיפו 100 μL של GAB לתערובת האקטין וערבבו היטב על ידי צנרת עדינה למעלה ולמטה. דגירה על קרח במשך שעה.
- הכינו APB טרי במהלך הדגירה.
- לאחר שלב הדגירה, פילמר את האקטין על ידי הוספת 11 μL של APB לתמיסת האקטין. מערבבים היטב על ידי צנרת למעלה ולמטה בעדינות. דגירה על קרח במשך 20 דקות.
- הוסיפו 5 מיקרון ליטר של פאלוידין עם תווית Alexa Fluor 488 ודגרו על קרח בחושך למשך שעה אחת.
- אחסנו את בקבוקון האקטין הביוטינילי העטוף ברדיד אלומיניום בחושך בטמפרטורה של 4 מעלות צלזיוס.
הערה: ניתן להשתמש בחוטים אלה לתקופה מקסימלית של שבוע אחד.
3. הכנת מיוזין וחרוזים
- שחזור מיוזין II
- סובבו לזמן קצר (~5 שניות) מיוזין שלד ליופילי II כדי לאסוף אותו בתחתית הצינור באמצעות מיניצנטריפוגה סטנדרטית.
- שחזרו את המיוזין ל-10 מ"ג/מ"ל על ידי הוספת 100 מיקרו-ליטר של 1 mM DTT שהוכן במי RO.
- דלל את תמיסת המיוזין 10x על ידי הוספת 10 μL של 10 מ"ג/מ"ל מיוזין ל-90 μL של 1 mM DTT במי RO. הפוך את הנפחים הקטנים (1-5 μL) לאליקוטים, הקפאה בהצמדה ואחסון בטמפרטורה של -80 °C.
הערה: ניתן לאשר את פעילות מיוזין על ידי ביצוע בדיקת נימה סטנדרטית כפי שפורסם בעבר46,47. ראה את הדיון לתיאור קצר.
- ניקוי חרוזים מצופים סטרפטאבידין
- יש לדלל 20 μL של חרוזי סטרפטווידין בנפח 1 מיקרומטר ל-80 μL של מי RO. יש לשטוף ארבע פעמים על ידי סיבוב כלפי מטה ב-9,600 × גרם ולשחזר את השימוש ב-100 מיקרו-ליטר של מי RO.
- סוניקט במשך 2 דקות באמפליטודה של 40% ואחסן את החרוזים השטופים על סיבוב ב-4 מעלות צלזיוס.
4. הכנת תאי זרימה
- הכינו תמיסת פולי-ל-ליזין (PLL) על ידי הוספת 30 מ"ל של 100% אתנול לצינור של 50 מ"ל והוספת 200 מיקרון של 0.1% פולי-ל-ליזין במים וערבבו היטב.
- הוסיפו מכסה חרוט לתמיסת ה-PLL ואפשרו לה להיספג למשך 15 דקות. הסירו את הכיסוי בעזרת פינצטה, והקפידו לגעת רק בקצה הכיסוי כשהוא נמשך כלפי מעלה מהצינור (ראו איור 1A-C). תפסו את הכיסויים בשוליהם ביד עם כפפות.
- יבשו את הכיסוי בחברת תעופה מסוננת עד שלא נותר אתנול ולא שאריות על הכיסוי.
- מרחו שתי חתיכות של סרט דביק דו-צדדי על אמצע שקופית מיקרוסקופ, במרחק של 3-4 מ"מ זו מזו. קרע או גזור את סרט ההדבקה העודף שתלוי מקצה המגלשה.
- הוסף את הכיסוי המצופה PLL על גבי סרט ההדבקה הניצב לציר הארוך של שקופית המיקרוסקופ (יצירת T) כדי ליצור ערוץ.
- השתמשו בצינור קטן כדי לדחוס את הכיסוי אל הסרט והמיקרוסקופ החליקו ביסודיות עד שהקלטת תהיה שקופה (איור 1A). ודא שאין בועות בקלטת מכיוון שהדבר עלול לגרום לדליפה מערוץ הזרימה.
הערה: תא הזרימה יכול להכיל נפח של 10-15 μL.
5. הכנת צרור אקטומיוסין
- בצינורות נפרדים, יש לדלל כל סוג של נימה אקטין (רודמין וביוטינילציה המסומנת 488) פי 600 על ידי ערבוב של 0.5 μL של האקטין המכובד, המסומן עם 300 μL של APB. הוסיפו 5 μL נוספים של הפלואידין המסומן בהתאמה לכל צינור ודגרו על קרח בחושך במשך 15 דקות.
- לתמיסת האקטין הביוטינילית, יש להוסיף מערכת לנטרול חמצן של 1 μL של בטא-D-גלוקוז ב-500 מ"ג/מ"ל, 1 μL של גלוקוז אוקסידאז ב-25 מ"ג/מ"ל, ו-1 μL של קטלאז ב-500 יחידות/מ"ל. הוסף 1 μL של 100 mM ATP ו 1 μL של 100x מדולל חרוזי streptavidin מנוקה. מערבבים בעדינות עם קצה פיפטה. שים את המתלה על סיבוב ב 4 מעלות צלזיוס בזמן שאר צרור actomyosin הוא להיות מורכב.
- הוסיפו 15 μL של רודמין אקטין מדולל לתא הזרימה PLL (איור 1D). לפתות את התמיסה העודפת דרך תא הזרימה אך לא לאפשר לתעלת הזרימה להתייבש. לדגור במשך 10 דקות בתא לחות.
הערה: ניתן ליצור תאי לחות מקופסאות קצה פיפטה ריקות עם תוספת מים לתחתית והמכסה מכוסה ברדיד אלומיניום כדי לחסום אור. - הכינו תמיסת קזאין של 1 מ"ג/מ"ל ב-APB.
- הוסיפו 15 μL של קזאין 1 מ"ג/מ"ל כדי למנוע קשירה לא ספציפית של הרכיבים הבאים (איור 1E). לדגור במשך 5 דקות בתא לחות.
- הוסף את הריכוז הרצוי של מיוזין לאקטין הביוטיניל ותרחיף החרוזים משלב 5.2. ערבבו בעדינות עם קצה הפיפטה, ולאחר מכן הוסיפו מיד 15 μL מתרחיף שלב 5.2 + ריכוז המיוזין הרצוי לתא הזרימה (איור 1F,G). דגירה במשך 20 דקות. אטמו את הקצוות הפתוחים של תא הזרימה עם לק כדי למנוע אידוי במהלך ניסויי הדמיה ולכידה אופטית.
הערה: ריכוז תמיסת מיוזין של 1 μM מניב קשירה חזקה וניתן להשתמש בה כנקודת מוצא להתאמה האישית הרצויה של הבדיקה (ראה איור 2).
6. מדידות כוח באמצעות מלכודת אופטית (NT2 Nanotracker2)
הערה: בעוד שהפרוטוקול שלהלן מיועד במיוחד למערכת NT2, ניתן להשתמש בבדיקה זו עם מערכות השמנה אופטיות אחרות, כולל אלה שנבנו בהתאמה אישית, שיש להן גם יכולות פלואורסצנטיות. זרימת העבודה הכללית נשארת זהה של מיקוד פני השטח של השקופית, ביצוע כיול חרוזים ורכישת נתונים על ידי מציאת חבילות אקטין פלואורסצנטיות. עבור מערכת NT2, איור משלים S1, איור משלים S2, איור משלים S3, איור משלים S4, איור משלים S5, איור משלים S6 ואיור משלים S7 מספקים פרטים על מערכת ההשמנה האופטית וממשק התוכנה.
- הפעל את תיבת הבקרה ואת הלייזר (איור משלים S1).
- הפעל את תוכנת מחשב המלכודת האופטית על ידי לחיצה על סמל JPK Nanotracker בשולחן העבודה.
- העירו את השלט הרחוק על ידי לחיצה על כפתור Logitech במרכז (איור משלים S2).
- הפעל את מודול הפלואורסצנציה על-ידי החלפת מתג ההפעלה/כיבוי (איור משלים S3).
- סובב את צריח קוביית המסנן להדמיית שדה בהיר (איור משלים S4).
- לאחר שהמערכת מוכנה, הפעל את הלייזר באמצעות לחצן הפעלת הלייזר בפינה השמאלית התחתונה של המסך ל-50 mW ותן לו להתייצב למשך 30 דקות (איור משלים S5).
- לחץ ברצף על כפתורי התאורה, המצלמה, המטרה והתנועה הבימתית בתוך התוכנה כדי להעלות חלונות אלה לצפייה ומניפולציה במהלך הניסוי. הפעל את תאורת המיקרוסקופ על-ידי לחיצה על לחצן ההפעלה/כיבוי והגדרתו לעוצמה מרבית על-ידי לחיצה וגרירה של הסרגל כל הדרך ימינה (איור משלים S5).
- פתח את אזור הדגימה והסר את מחזיק הדגימה משלב המיקרוסקופ. הוסף את תא הזרימה, אבטח אותו באמצעות מחזיקי דגימת המתכת, וודא שהשקופית עם הכיסוי נמצאת בתחתית.
- הוסף 30 μL של מי RO למרכז המטרה התחתונה. אין לתת לקצה הפיפטה לגעת בעדשה. הכנס מחדש את שלב הדוגמה.
הערה: מכיוון שמערכת NT2 משתמשת במטרה טבילה במים כמטרת ההשמנה, מדיית הטבילה עשויה להיות שונה בהתאם למטרת ההשמנה בהגדרת המשתמש. - הרם את המטרה הנמוכה יותר באמצעות חיצי הבקרה שעל המסך או L2 בשלט הרחוק עד שחרוז המים ייגע בכיסוי (איור משלים S5).
- הנמיכו את המטרה העליונה עד שתגיעו לכמחצית המרחק לתא הזרימה באמצעות החצים שעל המסך או R2 בשלט הרחוק. הוסף 170 μL של מי RO לחלק העליון של תא הזרימה ישירות מתחת למטרה העליונה. הנמיכו את המטרה העליונה עד שהיא שוברת את מתח הפנים של המים ויוצרת מניסקוס.
- הזז את שלב המיקרוסקופ באמצעות משטח החצים בשלט הרחוק עד שתגיע לקצה הקלטת הסמוכה לערוץ הזרימה. סגור את הדלת לדוגמה.
הערה: "לחיצה" בעת סגירת הדלת לדוגמה מציינת שתריס הלייזר פתוח כעת. זוהי תכונת בטיחות המאפשרת לתריס להיפתח רק אם הדלת סגורה. - באמצעות החלון ' מטרה' במסך, התמקד בקצה הקלטת על-ידי העלאת המטרה התחתונה בשם 'מטרת לייזר ' על-ידי לחיצה על החץ העליון באמצעות הפקדים שעל המסך. עשה את אותו הדבר עבור המטרה העליונה על-ידי לחיצה על החץ התחתון (איור משלים S5).
הערה: החצים הכפולים מזיזים את המטרה או את השלב מהר יותר. קצה הקלטת משמש למיקוד מכיוון שמדובר באובייקט גדול וקל למציאה הקרוב למשטח הכיסוי. בועות אוויר בתוך הקלטת הן אפשרות נוספת. עם זאת, הדבר אינו נדרש אם למשתמש יש שגרה אוטומטית למציאת מיקוד פני השטח או שיטה מועדפת בתוך החברה. - ברגע שהקלטת ממוקדת, סגרו חלקית את הקשתית בחלק העליון של המלכודת האופטית. הורידו את המטרה העליונה כלפי מטה עד שצורת המצולע של הקשתית נראית לעין. מקד את הקצוות האלה, פתח מחדש את הקשתית, ולאחר מכן הצמיד את המטרות באמצעות לחיצה על סמל המנעול (איור משלים S5).
- מצא חרוז צף ולכוד אותו על ידי לחיצה על כפתור תריס המלכודת , אשר יפתח את התריס ויאפשר ללייזר ההשמנה לפגוע בדגימה. לחץ על סמן ההשמנה על המסך וגרור אותו כדי להזיז את מיקום לייזר ההשמנה . לאחר הלכידה, כייל את החרוז כך שיתאים את מדידות המתח לכוח ולתזוזה.
- לחץ על כפתור הכיול . התאם את שגרת הכיול בהתבסס על ניתוח ספקטרום ההספק והתאם את תדר הפינה בתוך התוכנה לכיווני X, Y ו- Z (איור משלים S6).
- לחץ על הגדרות. הקלד את קוטר החרוז (1,000 ננומטר), והקלד את הטמפרטורה של השלב שנמצא בפינה השמאלית התחתונה של חלון התוכנה. (ראו איור משלים S6).
- לחץ על מלכודת 1. לחץ על אות X. לחץ על הפעל כדי לבצע את התאמת התדר הפינתי. לחץ וגרור בתוך החלון כדי למטב את התאמת הפונקציה. לחץ על השתמש בו לקבלת ערכי רגישות ונוקשות. לחץ על קבל ערכים. חזור על הפעולה עבור אותות Y ו- Z. סגור את החלון. (ראו איור משלים S6).
הערה: שגרות כיול חרוזים במערכות השמנה אופטיות אחרות או במערכות שנבנו בהתאמה אישית שנבדקו בחוזקה על ידי המשתמש, כגון שיטת שיווי המשקל או שיטת כוח הגרירה, מקובלות גםהן 57,58. - מצא צרור אקטומיוזין על ידי חיפוש חרוזים הקשורים ל- AFs על פני השטח של הכיסוי.
- כאשר מתגלה חרוז שאינו צפוף על ידי חרוזים צפים אחרים, התבונן ב- AFs סביבו על ידי הדמיה פלואורסצנטית כדי לאמת את נוכחותו של צרור.
- ודא שצרור קיים על-ידי חיפוש שני ה-AFs הפלואורסצנטיים. הפעל את מקור האור הלבן והשתמש בקוביית המסנן המתאימה כדי לדמות כל נימה של אקטין על ידי סיבוב הצריח (קוביות מסנן עירור של 488 ננומטר ו- 532 ננומטר עבור אלקסה פלואור 488 ועירור רודמין, בהתאמה). ראו איור משלים S4.
הערה: ניסוי בקרה לאימות עוצמת הפלואורסצנטיות של AFs בודדים יכול להיות שימושי בזיהוי חבילות המורכבות מחוט יחיד עם תווית של 488 ו-rhodamine, או ישים לכל קבוצה של פלואורופורים שהמשתמש בוחר להשתמש בה. - לאחר האימות, לכוד את החרוז המחובר לנמה העליונה של החבילה על ידי לחיצה על לחצן תריס מלכודת .
- השתמש בפקדים שעל המסך כדי להקליט את הנתונים על-ידי לחיצה על לחצן אוסצילוסקופ (איור משלים S7). כדי להציג באופן חזותי את המדידות מבלי להקליט את הנתונים, לחץ על התחל. כדי לשמור את כל הנתונים, לחץ על שמירה אוטומטית. כדי להקליט מדידות, לחץ על התחל הקלטה. בחר אילו נתונים יש להציג באופן חזותי בזמן אמת (מיקום, כוח, כיוון x, כיוון y) על ידי בחירה מתוך התפריט הנפתח אות X או אות Y. זכור כי xdirection הוא משמאל לימין, וכיוון y הוא למעלה ולמטה על המסך. ראו איור משלים S7.
הערה: הנתונים יישמרו כקבצי .out וכוללים זמן, מתח, תזוזה וכוח לכל כיוון. ניתן לייצא קבצים אלה לתוכנות אחרות לצורך תצוגה חזותית וניתוח.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
תאי זרימה המכילים את מערכות צרור האקטומיוזין הם בעלי עיצוב סטנדרטי, המורכב משקופית מיקרוסקופ וכיסוי חרוט המופרדים על ידי תעלה העשויה מסרט דביק דו-צדדי (איור 1). לאחר מכן, הבדיקה נבנית מהכיסוי באמצעות מבואות מבוימים כמתואר בפרוטוקול. הבדיקה הסופית מורכבת מחוטים של אקטין המסומנים ברודמין; ריכוז המיוזין הרצוי (1 μM שימש לתוצאות המייצגות באיור 2 ובאיור 3); ביוטינילציה, אלקסה פלואור 488 עם תווית חוטי אקטין; 1 מיקרומטר חרוזי סטרפטאווידין; מערכת טיהור החמצן; ATP; ומאגר APB. צרורות מרובים ייווצרו לכל תא זרימה, וריכוזי האקטין שתוארו לעיל נותנים ריווח הולם בין צרורות כדי להבטיח שלא יהיו אינטראקציות לא רצויות. זה גם מקל על קבלת מדידות כוח מרובות לכל תא זרימה כדי להגביר את יעילות רכישת הנתונים. פרופילי כוח צריכים להיות ניתנים לשחזור בתוך תא זרימה ומתא זרימה לתא זרימה.
בעוד שהפרוטוקול לעיל מכוון לשימוש במערך השמנה אופטי מסחרי, תא הזרימה והבדיקה המוצגים כאן יכולים לשמש בקלות למכשיר מסחרי אחר או למערך השמנה אופטי שנבנה בהתאמה אישית בשילוב עם שלב מיקרוסקופ או מיקרוסקופ ובעל יכולות הדמיה פלואורסצנטיות. לאחר השלמת כל התוספות של תאי הזרימה בהתאם לפרוטוקול לעיל, צרורות האקטומיוזין בשקופית (איור 1) מוכנים למדידה מיידית. תא הזרימה מתווסף לשלב מיקרוסקופ המלכודת האופטית, מדידות כיול חרוזים מרובות נרכשות, וחבילות מזוהות באמצעות קולוקליזציה פלואורסצנטית של חוטי הצרור. חרוז הקשור לצרור נלכד, והתזוזה ומדידת הכוח המתאימה מתחילה. המשתמש יכול לצפות ברכישת נתונים בזמן אמת על צג המחשב. בהתאם לריכוז המיוזין המשמש בתא הזרימה, הצרור יכול להתחיל להפגין תנועה משמעותית באופן מיידי, או שייקח 30 s-1 דקה כדי לראות ביעילות עלייה בתזוזה / כוח.
עקבות כוח מייצגים מוצגים באיור 3A שבו מנועי המיוזין מפגינים רמפה יציבה בכוח ואחריה מישור. זה אופייני לראות סוגים אלה של עקבות להתפתח במשך 2-5 דקות. עם זאת, ניתן גם למדוד צרורות אקטומיוזין שאינם מייצרים כוח נטו כלשהו (איור 3B). עקבות אלה מופיעים כרעש בסיסי או אינם מציגים עלייה משמעותית נטו בכוח מעל 90 שניות. זה כנראה בגלל ריכוז מקומי נמוך של מנוע שאינו מאפשר החלקה פרודוקטיבית, או שהצרור נמצא בכיוון מקבילי שלילי שבו קצוות הפלוס והמינוס של החוטים מיושרים.
מכיוון שתכולת תא הזרימה עלולה להיות רגישה להתפרקות כתוצאה מתאורת האירוע ולייזר הלכידה, חימום מקומי במגלשה לאורך זמן ויצירת מיני חמצן רדיקליים, מומלץ מאוד לא להשתמש באותו תא זרימה במשך יותר משעה. ליעילות מרבית, מוצע לבצע בדיקה נוספת תוך כדי רכישת נתונים. ניתן לייצא מעקב תזוזה/כוח מתוכנת ההשמנה האופטית ל- Excel, Matlab, Igor או תוכניות אחרות לניהול נתונים לצורך סינון וניתוח נוספים. נתונים שניתן לחלץ מניסויי אנסמבל/צרור אופטיים כאלה כוללים סוגים שונים של פרופילי יצירת כוח (קו בסיס, רמפה/מישור) בתנאי בדיקה משתנים, מהירות יצירת כוח, יצירת כוח מרבית, התנהגות קינטית ודריכה של אנסמבל באמצעות גדלי צעדים וזמני שהייה בין צעדים או צוותי צעדים, כמו גם יחס חובה. המשתמש יכול גם לשנות את תנאי הבדיקה כדי להשוות כיצד הוספת סוגים שונים של מנועי מיוזין, הוספת חלבונים קושרי אקטין או שינוי תנאי חיץ משפיעים על מאפייני יצירת כוח אנסמבל אלה.
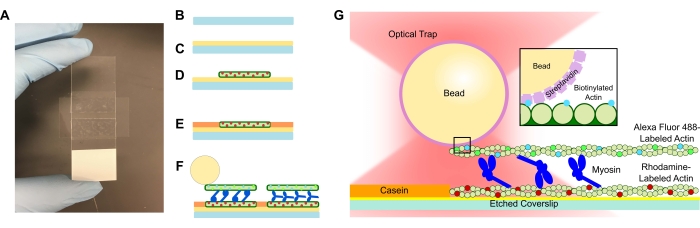
איור 1: בדיקה סכמטית. (A-C) כיסויים חרוטים מצופים בפולי-L-ליזין ומשמשים ליצירת תא הזרימה באמצעות סרט דו-צדדי ושקופית מיקרוסקופ. מבואות מתוזמנים ושלבי דגירה המתוארים בפרוטוקול גורמים לכך שאקטין מיוצב פאלוידין המסומן ברודמין הוא התבנית או החוט התחתון (D), ולאחר מכן חסימת קזאין למניעת קשירה לא ספציפית (E), ו-(F) Alexa Fluor 488 אקטין מיוצב פלואידין כמטען או נימה עליונה, וצוותים של מיוזין II שמחליקים את החוטים זה מזה ומייצרים כוח כאשר ATP מוצג. הגיאומטריה של המנועים ואופי ההצלבה בתוך הצרור עשויים להשתנות בתנאים שונים, כגון ריכוז מלח59. מחקרים קודמים הראו כי לתחום זנב המיוזין יש את היכולת לתקשר עם חוטי אקטין ותנועתיות אנסמבל איטית46. עם זאת, ראשי מיוזין בניסויי מרומיוזין כבדים מדגימים קשירה של כל ראש לחוטים סמוכים של אקטין60. (G) חרוזי סטרפטווידין משמשים כידית האופטית של המלכודת ונקשרים אך ורק לנימה של אקטין ביוטיניל, מה שמסייע לאמת כי צרורות תקינים נוצרים על המגלשה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.

איור 2: צרורות אקטומיוזין פלואורסצנטיים. ארבעה מפגשים שונים של חוטי אקטין וצרורות בתוך מבחן הצרור המוצגים באיור 1. נימת האקטין הביוטינילית העליונה עם תעלת הפלואור 488 פאלוידין של Alexa מוצגת משמאל, ונימת האקטין בתבנית התחתונה עם תעלת הפלואידין רודמין נמצאת בצד ימין. בחלק התחתון, אותה דמות מוצגת עם קווים צבעוניים מעל כדי לעזור להנחות את העין. (A) נימה של אקטין עליון נמצאת ליד נימה של אקטין תחתון אך יש לה כיסוי לא שלם. זה לא ישמש לניסויים צרורים. (B) חוטי אקטין עליונים ותחתונים הם קולוקליים, ועוצמת כל נימה מאשרת שכל אחד מהם הוא חוטים בודדים בתוך הצרור. זה יהיה מועמד טוב לניסויים צרורים. (C) צרור גדול של חוטי רודמין בהרכבה עצמית נמצא בתחתית. אמנם יש נימה אקטין עליונה מקבילה שהיא קולוקלית, אך יש יותר מדי חוטים תחתונים; לפיכך, הוא לא ישמש לניסויים צרורים. זוהי גם דוגמה לאופן שבו כאשר חוטי אקטין מרובים מאותו סוג כרוכים יחד, עוצמת הפלואורסצנטיות עולה. המשתמש יכול להשתמש בזה כמד לשיפוט חוטים בודדים לעומת חבילות מאותו סוג נימה. (D) נימה תחתונה קיימת ללא נימה עליונה תואמת, וגם היא אינה מאשרת דימום. זה לא ישמש לניסויים צרורים. נציין כי עוצמת החוטים בערוץ אלקסה פלואור 488 נמוכה ומאמינים שהיא נובעת מסט המסננים בו נעשה שימוש (סט פילטרים 09 מ- Zeiss). ערכת המסננים המשמשת לתעלת הרודמין היא ערכת מסננים 43 מבית Zeiss. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.

איור 3: יצירת כוח מאנסמבל מיוזין II. עקבות מייצגים של מנועי מיוזין II שלדיים המייצרים כוח בתוך ההיררכיה המבנית של אקטין במבחנה . מנועי המיוזין עובדים יחד כדי לייצר כוח באופן קולקטיבי ופרודוקטיבי עד שמגיעים לרמה והכוח מתקיים (A) או חווים אנטגוניזם ליד קו הבסיס (B). אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
איור משלים S1: מלכודת אופטית של Bruker/JPK Nanotracker2. (A) צג מחשב. (B) מקלדת מחשב. (C) מגדל מחשב. (ד) תיבת בקר. (E) ספק כוח לייזר. (F) תיבת אופטיקה של השמנה אופטית. (G) מיקרוסקופ הפוך. (H) שלב מהדלת למיקרוסקופ. (I) מחוון מקטב למעבר בין הדמיית ניגודיות של הפרעות בהירות ודיפרנציאליות. אנא לחץ כאן כדי להוריד קובץ זה.
איור משלים S2: שלט רחוק למלכודת אופטית. (A) לוח מקשים למיקום השלב הממונע. (ב-ג) התאם את מיקום ההשמנה. (D) A, X ו-B מפעילים ומכבים את התריס הראשי, את תריס המלכודת 1 ואת תריס ההשמנה 2, בהתאמה. (E) לחצן Logitech משמש להעיר את הבקר. (F) הלחצנים למעלה ולמטה המשמשים למיקום מטרת ההשמנה. (G) הלחצנים למעלה ולמטה המשמשים למיקום מטרת הזיהוי. שים לב שהשלט הרחוק אינו נדרש, וניתן לבצע את כל המניפולציות הללו בתוכנה. עם זאת, נוח להיות מסוגל לשלוט על המטרות ועל מיקום הבמה תוך הסתכלות לתוך סביבת הבמה מיקרוסקופ. אנא לחץ כאן כדי להוריד קובץ זה.
איור משלים S3: מודול פלואורסצנציה למלכודת אופטית. מקור האור הלבן הפלואורסצנטי של 89North PhotoFluor מוצמד לחלק האחורי של המיקרוסקופ ההפוך. הוא מופעל וכבה באמצעות מתג דו-מצבי (חץ). אנא לחץ כאן כדי להוריד קובץ זה.
איור משלים S4: צריח קוביית מסנן פלואורסצנטי. ניתן להפוך את הצריח (חץ) כדי להשתמש בקוביית המסנן הדרושה להדמיה בצבעי DIC, רודמין או אלקסה פלואור 488. שים לב שניתן להחליף קוביות מסנן כדי להתאים אישית את ההגדרה לשימוש בפלואורופורים שונים. אנא לחץ כאן כדי להוריד קובץ זה.
איור משלים S5: תוכנת Nanotracker2. (A) לחצן הפעלה לייזר ובקרה. (B) חלון מיקום אובייקטיבי. חצים כיווניים משמשים להזזת מטרות הזיהוי (למעלה) וההשמנה (למטה). חצים כפולים מזיזים את המטרות במהירות גבוהה יותר. הכפתור הכחול והאדום בפינה השמאלית התחתונה מבטל את הזוגות של המטרות ומחזיר אותן למקומן המקורי. זה הכרחי בעת לקיחת דגימות בשלב המיקרוסקופ ומחוצה לו. הכפתור השלישי משמאל עם המטרות וסמל המנעול "מצמיד" את המטרות כך שכאשר הן נמצאות בפוקוס ומשיגות תאורת קולר, המשתמש יכול להזיז את מטרות ההשמנה והזיהוי למעלה ולמטה בציר z. (C) חלון מיקום לדוגמה המשמש להזזת שלב המיקרוסקופ בציר x ו-y. חצים כפולים מזיזים את הבמה במהירות גבוהה יותר. חלון זה מופעל על ידי לחיצה על סמל החץ למעלה / למטה ושמאלה / ימינה בתפריט העליון. (D) חלון הדמיה של מצלמה. ניתן להשתמש בסמל מפתח הברגים כדי להגדיר תנאי הדמיה מותאמים אישית. חלון זה מופעל על ידי לחיצה על מצלמה סמל בתפריט העליון. (E) חלון תאורת מיקרוסקופ. חלון זה מופעל על ידי לחיצה על סמל הנורה בתפריט העליון. אנא לחץ כאן כדי להוריד קובץ זה.
איור משלים S6: חלון כיול. (א) חלון זה משמש לכיול חרוזים ומופעל על ידי לחיצה על סמל Cal בתפריט העליון. כדי לכייל חרוז, ההתאמה הטובה ביותר של תדר הפינה מתבצעת באותות x, y ו- z. (B) עבור כל אות, בחר את לחצן האות המתאים בפינה השמאלית העליונה. (C) לחץ על הפעלה ומטב את ההתאמה על ידי לחיצה וגרירה בתוך החלון הירוק (D). (E) לאחר שתהיו מרוצים מההתאמה, לחצו על 'השתמש בו ' לרגישות ונוקשות. זה יאפשר רישום תזוזה בננומטרים וכוח בפיקונווטונים. (F) לאחר מכן, לחץ על קבל ערכים בפינה השמאלית התחתונה. חזור על הפעולה לקבלת הכיוונים y ו- z. אנא לחץ כאן כדי להוריד קובץ זה.
איור משלים S7: חלון רכישת נתונים. חלון זה משמש לרכישת נתוני מיקום וכוח ומאפשר למשתמש לראות את המדידות בזמן אמת. (A) חלון זה מופעל על ידי לחיצה על סמל x,t בתפריט העליון. (B) המשתמש יכול לעבור בין צפייה באותות x ו- y. (C) לחץ על התחל כדי להתחיל להציג נתונים באופן חזותי. לחץ על שמירה אוטומטית כדי לשמור את הנתונים. לחץ על התחל להקליט כדי להתחיל להקליט ולשמור נתונים. אנא לחץ כאן כדי להוריד קובץ זה.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
מחקר במבחנה באמצעות פינצטה אופטית בשילוב עם הדמיה פלואורסצנטית בוצע כדי לחקור את הדינמיקה של הרכבי מיוזין המקיימים אינטראקציה עם חוטי אקטין. צרורות אקטין-מיוזין-אקטין הורכבו באמצעות מיוזין שריר II, רודמין אקטין בתחתית הצרור ועל משטח הכיסוי, וסיבי אקטין בעלי תווית של 488 ביוטינילים בחלק העליון של הצרור. חלבון אקטין משרירי ארנב עבר פולימריזציה וייצוב באמצעות מאגרי אקטין כלליים (GAB) ומאגרי פילמור אקטין (APB). GAB ו-APB חייבים להיות מוכנים טריים מדי יום במעבדה באמצעות ATP, מאגר FC ומאגר TC. מיוזין שריר II שימש ליצירת כריכי אקטין-מיוזין-אקטין. פלואידין שימש להכתמה פלואורסצנטית של חוטי האטין, כמו גם לייצוב במבחנה.
ניתן לאשר את פעילות מיוזין על ידי ביצוע בדיקת נימה סטנדרטית כפי שפורסם בעבר46,47. מיוזין II ותת-החלקיקים שלו יכולים להיקשר למשטח הכיסוי במגוון כיוונים, והנוכחות של תחום הזנב יכולה להאט את החלקת החוטים בהשוואה למבחנים המשתמשים במרומיוזין כבד46,48,49. עם זאת, עדיין ניתן להבחין בגלישה ובתנועת פני השטח. הדגמה בולטת יותר של פעילות מיוזין היא שבירת נימה אקטיבית של אקטין שניתן להבחין בה כאשר חוטי אקטין ארוכים יותר נשברים לרסיסים קטנים יותר שמחליקים משם לכיוונים מרובים. זה קורה בשל הריכוז הגבוה של מנועים פעילים על פני השטח, נצפה על ידי מספר מעבדות, ואינו מתרחש ללא מנועי מיוזין פעילים נוכחים 42,50,51,52,53,54. יתר על כן, מבחן הצרור המוצג כאן מסייע בהקלה על בעיות תנועתיות שהיו קשורות בעיקר לבדיקת חוט הגלישה, כגון מגוון כיווני קשירת המנוע על מכסה זכוכית, מכיוון שבדיקת הצרור כוללת חסימת קזאין של משטח הזכוכית כך שהמנועים נקשרים בתוך הצרור 47,55,56.
הצעד הראשון הוא להוסיף חוטי רודמין אקטין כחוט ההימה התחתון או התבנית לכיסוי מצופה פולי-L-ליזין בתא זרימה. פולי-ל-ליזין משמש לקידום קשירת אקטין מכיוון שפולי-ליזין טעון חיובית ואילו לאקטין יש מטענים שליליים והוא שימש בתכשירי בדיקה ציטוסקטליים קודמים במבחנה 61,62,63. לפני היווצרות הצרור, נוספו דילולי אקטין שונים לתא זרימה כדי לייעל את ריכוז האטין. במקרה זה, 600x מהמלאי היה הדילול האופטימלי שהניב מספר מספיק של חוטי תבנית ליצירת צרור אך עם ריווח הולם כך שהצרורות היו מותאמים אישית. הדילול בוצע באמצעות חיץ APB. הוספת רודמין אקטין לוותה בשכבת קזאין כדי לחסום את פני השטח ולמנוע קשירה לא ספציפית. תא הזרימה דוגר במשך 30 דקות ונשטף לאחר הדגירה עם חיץ כדי לשטוף את כל חוטי האקטין הלא מאוגדים. לבסוף, שילוב של מיוזין, 488/ביוטין אקטין וחרוזים מצופים סטרפטווידין נוספו לתא הזרימה כדי להקל על היווצרות צרור אקטין-מיוזין. ריכוז החרוזים צריך להיות כזה שיש מספיק כדי לקשור צרורות הקשורים לפני השטח ומספיק בהשעיה כדי להקל על הכיול. עם זאת, ריכוז חרוזים גבוה מדי עלול לגרום לקושי במהלך ניסויי לכידה עקב נפילת חרוזים שכנים למלכודת הלייזר ושיבוש המדידה. מנועי מיוזין מתווספים לשילוב ממש לפני הזרקתו לשקופית, כך שמנועי המיוזין אינם מצטברים מראש עם המטען או עם נימה ביוטינילית עליונה ובכך יקשרו את הרודמין התחתון לצרור חוטי אקטין ביוטינילציה.
מערכת ההשמנה האופטית NT2 היא מלכודת אופטית מסחרית עם שילוב של שדה בהיר, ניגודיות הפרעה דיפרנציאלית (DIC) ושיטות הדמיה אפיפלואורסצנטיות. הוא משולב עם מיקרוסקופ הפוך Zeiss AxioObserver 3 עם מטרות לכידה וזיהוי מים של 100x/NA 1.46 ו-63x/NA 1.0. המערכת מצוידת ביכולת השמנה של קליק וגרירה של מלכודת לייזר אחת וניתן להשתמש בה בעת הדמיה בכל אחת מהשיטות המפורטות בעבר. הצרורות שנוצרו מזוהים ומאושרים באמצעות הדמיה פלואורסצנטית. מקור אור לבן עם קוביות סינון מתאימות (GFP/FITC ו-TRITC/CY3) מאפשר מעבר מהיר בין הדמיית נימה. AFs שעברו Colocalized אומתו על ידי הדמיה של ה-AFs באורכי גל העירור השונים לפני ביצוע כל מדידת כוח באמצעות פינצטה אופטית. מכיוון שהחוטים יכולים לבצע פוטו-אקונומיקה במהירות אפילו עם מגיב מנטרל חמצן, מוצע לחוקרים לייעל פרמטרים של הדמיה כגון עוצמה וזמן חשיפה לפני ביצוע ניסויי הצרור.
לכידה אופטית שימשה לביצוע מדידות הכוח, תוך שימוש בחרוזי הסטרפטווידין בנוכחות ATP כדי לקשור את נימה אקטין המטען הביוטינילי ולהפעיל את ייצור כוח המיוזין כמתמר כוח. נתוני תזוזה וכוח לעומת זמן שהתקבלו על ידי השמנה אופטית חולצו מתוכנת ההשמנה לצורך ניתוח. עם זאת, תוכנת ההשמנה המסחרית מספקת גם שגרות ניתוח שניתן להשתמש בהן, או שאלגוריתמים מותאמים אישית בתוכניות אחרות יכולים להיות מתוכנתים על ידי המשתמש כדי להמחיש ולנתח נתוני השמנה. במערכות השמנה אופטיות מותאמות אישית, המשתמש עשוי להיות לייזר עירור במקום מקור אור לבן עם מסננים, שגם הם מקובלים לשימוש. יתר על כן, ניתן לשנות צבעים פלואורסצנטיים כך שיתאימו לציוד הקיים שיש למשתמש אם ספקטרום הפליטה אינו חופף וגורם לדימום.
נציין כי הבדיקה המוצגת היא בדיקה בסיסית הניתנת להתאמה אישית נוספת על ידי המשתמש בהתאם לשאלת המחקר שלו בתחום מכניקת האנסמבל actomyosin. ניתן ליישם את זרימת העבודה הכללית גם על מערכות אנסמבל ציטוסקטליות אחרות במבחנה שעשויות לעניין, כגון מבחני צרור מיקרוטובולים היוצרים מודלים מינימליים של ציר מיטוטי 32,61,63,64,65,66. שינויים יכולים לכלול, בין היתר, שינוי תוויות הפלואורופור המתאימות להגדרה הקיימת של המשתמש; שינוי ריכוז מיוזין, מבנה או איזוטיפ; ותנאי חיץ טיטרציה, בין היתר.
אתגרים פוטנציאליים אפשריים בעת ביצוע בדיקה זו. בעת יצירת צרורות אקטין-מיוזין, ריכוז המיוזין בתוך צרורות האקטין עשוי שלא להיות הומוגני לאורך השקופית. כדי להתאים לכך, יימדדו חבילות מרובות לאורך כל השקופית כדי להבטיח שפרופילי חלוקת המנוע וייצור הכוח יידגמו כראוי. זה גם מאתגר לדעת כיוון צרור אם זה נדרש לפרשנות של נתוני כוח. לכן, יש לקחת מספר ניסויים עבור כל צרור. אפשר גם לשלב תיוג קצה של נימה אקטין באמצעות ג'לסולין פלואורסצנטי או חרוזים מצופים ג'לסולין בגודל קטן יותר מאשר ידית ההשמנה האופטית. ניתן להשתמש בהדמיה פלואורסצנטית גם כדי לבחון את כוחות הרכיבים x ו- y כדי להסיק את כיוון הצרור. יתר על כן, מכיוון שמצב צבירת המיוזין מושפע מאוד מהחוזק היוני של החיץ עם היווצרות חוטים עבים המתרחשים עם דילול מהיר של KCl, ריכוז מלח החיץ צריך להיות מנוטר כראוי67,68.
מחקרים קודמים שהשתמשו בשיטות אחרות במבחנה, כגון מבחני גלישה, סייעו לזהות את תפקידם של תחומי המיוזין ולחקור את התצורה והאינטראקציות בין מיוזין לחלבונים קושרי אקטין אחרים. עם זאת, לשיטות אלה יש חיסרון בכך שקשירת מיוזין על משטח קשיח תגביל את פוטנציאל התיאום בין מנועי המיוזין ובכך תבצע משוב מכני המתרחש כדי לקבוע אם ההרכב המוטורי נמצא במצב יחס חובה גבוה או נמוך 33,35,41,69. יתר על כן, לכידה אופטית עם רשתות מנוע מיוזין יחיד אינה נותנת הבנה ברורה של האופן שבו מנועי מיוזין מתקשרים זה עם זה ועם חוטי אקטין. הפרוטוקול שפותח כאן מאפשר לחקור את הדינמיקה של אנסמבל מוטורי מיוזין בתוך רשת אקטין תואמת והיררכית. הוא גם ניתן להתאמה אישית במונחים של מאפייני אנסמבל נימה מוטורית כגון ריכוז, איזופורם וסביבת חיץ, בין היתר, כדי לאפשר חקירה שיטתית. הפרוטוקול המוצג הוא פלטפורמה למחקרים עתידיים של רשתות אקטומיוזין מורכבות יותר ושומר על הדיוק של מדידות תזוזה וייצור כוח המתאפשרות על ידי השמנה אופטית ששימשה באופן מסורתי למחקרים של מולקולות בודדות.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
למחברים אין ניגודי עניינים להצהיר.
Acknowledgments
עבודה זו נתמכת בחלקה על ידי מלגת המחקר של מועצת הסטודנטים לתארים מתקדמים של אוניברסיטת מיסיסיפי (OA), מכללת המצטיינים של אוניברסיטת מיסיסיפי סאלי מקדונל-בארקסדייל (JCW, JER), קונסורציום מענקי החלל של מיסיסיפי תחת מענק מספר NNX15AH78H (JCW, DNR), ואיגוד הלב האמריקאי תחת מענק מספר 848586 (DNR).
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
