

Summary
in vitroでのアクトミオシンバンドルの形成と光ピンセットを用いたミオシンアンサンブル力発生の測定について紹介し、議論します。
Abstract
ミオシンは、ATPを加水分解してアクチンフィラメント(AF)トラックに沿ってステップするモータータンパク質であり、運動性や筋肉収縮などの細胞プロセスに不可欠です。ミオシンIIは、その力発生機構を理解するために、1分子(SM)レベルと、光トラップなどの生物物理学的手法を用いた in vitro でのモーターチームとして研究されています。
これらの研究は、ミオシンの力発生挙動が、3ビーズ配置の単一分子レベルから、滑走配置の剛ビーズまたはカバーガラス表面上で一緒に動作するモーターのグループに移動すると大きく異なる可能性があることを示しました。ただし、これらのアッセイ構造では、細胞内のように粘弾性構造階層内のミオシンのグループダイナミクスを評価することはできません。我々は、光ピンセットを用いて、複数のアクチンフィラメントと相互作用するミオシンアンサンブルによる力発生のメカニズムを調べる方法を開発しました。
これらのアクトミオシンバンドルは、運動通信とアンサンブル力出力をキャプチャする階層的で準拠した環境での研究を容易にします。アッセイのカスタマイズ可能な性質により、実験条件を変更して、ミオシンアンサンブル、アクチンフィラメントバンドル、または周囲の環境への変更がどのようにして異なる力出力をもたらすかを理解することができます。
Introduction
モータータンパク質は生命に不可欠であり、化学エネルギーを機械的仕事に変換します1,2,3。ミオシンモーターは、フィラメントに沿ってトラックのようなステップを踏むことによってアクチンフィラメントと相互作用し、アクチン-ミオシンネットワークのダイナミクスは、筋肉の収縮、細胞の運動性、細胞質分裂中の収縮リング、および細胞内の貨物の移動など、重要なタスクを実行します3,4,5,6,7,8 .ミオシンは非常に多くの重要な役割を担っているため、ミオシン-アクチンネットワークの機能が機能しなくなると、肥大型心筋症(HCM)で心臓の過収縮を引き起こすミオシン重鎖の変異などの疾患発症につながる可能性があります9,10,11,12,13,14。.筋収縮において、個々のミオシンモーターは、AF4、15、16、17、18の相対的なスライドを実行する必要な機械的エネルギーを提供するためにアンサンブルとして働くことによって互いに協力する。ミオシンモーターは、AF間にクロスブリッジを形成し、そのメカノケミカルサイクルによる立体配座変化を利用して、整列したフィラメント17、18、19、20、21の有刺鉄線の端に向かって集合的に移動する。
光学トラップなどの技術を用いたSMレベルでの定量的in vitro運動アッセイの開発により、SM力の発生とステップサイズの測定など、個々のミオシンモーターがどのように機能するかについての前例のない詳細を収集することが容易になりました22,23,24,25,26,27,28,29,30.Fineerらは、単一のミオシンIIモーターの力発生メカニズムを調べるために、「スリービーズ」または「ダンベル」光学トラップアッセイを開発しました23,31。筋ミオシンIIはチームでAFを収縮させるが、SMレベルでは非処理的であるため、光学トラップアッセイの向きは、古典的なモーター結合ビーズアプローチ32から再配置する必要がありました。ダンベルアッセイを形成するために、2つの光学トラップを使用して、カバーガラスに取り付けられたビーズに結合したミオシンモーター上にAFを保持し、トラップ23内のAFの動きによって単一のモーターによる力出力を測定した。
しかし、ミオシンIIを含む多くのモータータンパク質は単独で機能せず、しばしばそれらの部分の合計として機能しないため、SM力および単一モーター/単一フィラメントアッセイの向きを使用しても、システムレベルの力の生成に関する完全なイメージが得られません15,16,17,32,33,34,35,36 .ミオシンとアクチンフィラメントのネットワークの相乗効果をよりよく理解するには、複数のフィラメントと相互作用する複数のモーターを含むより複雑な構造が必要です15,32。ダンベルアッセイの配向は、ビーズに複数のミオシンを付着させるか、または表面に付着したミオシン太いフィラメントを使用して、モーターが懸濁AFと相互作用できるようにすることにより、小さなアンサンブル力の発生を調べるために利用されています4,23,34,37,38,39,40。
他の小さなアンサンブルアッセイには、ミオシンモーターがカバーガラス表面にコーティングされ、AFに結合したビーズを使用してモーターのチームによって生成された力をプローブするin vitroフィラメントグライディングアッセイが含まれます4,35,36,38,39,40,41,42,43.どちらの場合も、ミオシンは硬い表面(ビーズまたはカバーガラス)に結合し、1つのAFを利用します。これらの場合、モーターは自由に動くことも互いに通信することもできず、ミオシンをしっかりと束縛することは、モーターがサルコメア32で一緒に働くであろう準拠した階層的環境を反映していない。以前の研究では、ミオシンIIがその環境を感知し、力の発生やデューティ比などの特性を変更することにより、粘弾性またはモーター濃度条件の変化に応じて適応できることが示唆されています41,44,45。したがって、ミオシンIIアンサンブル力発生の機構的基盤のより現実的な絵を描くために、運動通信とシステムのコンプライアンスを促進およびキャプチャする光トラップアッセイを開発する必要があります。
ここでは、2本のアクチンフィラメント間で相互作用する複数のミオシンモーターからなるアクトミオシンバンドルまたはサンドイッチを形成することにより、in vitroで階層構造を光トラップと結合させる方法を開発しました。このモジュラーアッセイ形状は、分子的および環境的要因がアンサンブルミオシン力の生成にどのように影響するかを直接調査する機能を備えています。さらに、これらのアクチン-ミオシンアンサンブルを介した力発生メカニズムの調査は、筋肉収縮などの大規模な細胞タスクが分子レベルからどのように伝播するかをモデル化および理解するのに役立つ可能性があります9,10,13。
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1.エッチングカバーガラス
- 100 gのKOHを1,000 mLのビーカーに300 mLの100%エタノールに溶かします。KOHの大部分が溶解するまで攪拌子で攪拌します。
注意: 濃縮KOH溶液は、火傷や衣服の損傷を引き起こす可能性があります。手袋、目の保護具、白衣を着用してください。 - カバースリップをカバースリップクリーニングラックに個別に配置します。
注意: ラックは、カバースリップの各面のエッチングとすすぎを可能にするために間隔を空けて単一のカバースリップを保持するスリットで設計されており、底部の排水穴があり、過酷なエッチング条件に耐えることができる材料で作られています。それらはカスタムメイドまたは商業的に購入することができます。 - 3つの1,000 mLビーカーを準備してラベルを付けます:1つは300 mLのエタノールで、2つのビーカーは300 mLの逆浸透(RO)水で。
注:ここでは、RO水は実験室の浄水器から供給されましたが、地元の浄水器が利用できない場合は市販品を購入することもできます。 - 4つのビーカーのそれぞれをバス超音波処理器に入れて、5分間脱気します。
- KOHとエタノールのビーカーにカバーガラスのラックを沈め、5分間超音波処理します。
- カバーガラスのラックをKOH /エタノールビーカーからエタノールのみのビーカーに移します。ビードがなくなるまでビーカーでラックを上下に浸します。
注意: カバーガラスを乱したり、ラックをビーカーに無理に落としたりしないように注意してください。これにより、カバーガラスがラックから出たり、化学物質の飛沫が発生したりします。 - カバーガラスのラックをエタノールビーカーから水のビーカーに慎重に移し、ビーズがなくなるまで上下に浸漬します。
- まだ使用されていない水のビーカーにカバーガラスのラックを沈め、5分間再び超音波処理します。
- ボトルを使用して、カバースリップのラックにカバースリップからスムーズに流れ落ちるまで水をスプレーします。エタノールで繰り返します。
- ラックを90°Cのオーブンで20分間乾燥させます。エッチングされたカバーガラスのラックは、使用前に汚染を防ぐために、密閉容器に室温で保管してください。
2.アクチンフィラメント重合
- ソリューションTを作る
- 50 mLのコニカルチューブに、3.94 gのトリス塩酸塩と0.147 gのCaCl2を加えます。RO水を加えて総容量50mLにし、よく混ぜます。
注:溶液Tの最終濃度は、それぞれ500 mM Tris-HClと20 mM CaCl2 です。 - チューブ溶液Tにラベルを付け、4°Cで保管します。
- 50 mLのコニカルチューブに、3.94 gのトリス塩酸塩と0.147 gのCaCl2を加えます。RO水を加えて総容量50mLにし、よく混ぜます。
- TCバッファの作成
- 40 mLのRO水と1.5 mLの溶液Tを50 mLのコニカルチューブに混ぜます。少量の濃縮KOHを加えてpHを8.0に変更します。水を加えて50mLの溶液を作り、pHを確認します。必要に応じてpHを調整します。
注:最終的なTCバッファーには、pH 8で5 mM Tris-HClと0.2 mM CaCl2 が含まれています。 - チューブTCにラベルを付け、4°Cで保管します。
- 40 mLのRO水と1.5 mLの溶液Tを50 mLのコニカルチューブに混ぜます。少量の濃縮KOHを加えてpHを8.0に変更します。水を加えて50mLの溶液を作り、pHを確認します。必要に応じてpHを調整します。
- FCバッファの作成
- 85 mLのRO水、10 mLの溶液T、3.73 gのKCl、および0.041 gのMgCl2 を100 mLバッファーボトルに加えます。少量の濃縮KOHを加えてpHを7.5に変更します。水を加えて最終容量を100mLにし、pHを確認します。
注: 最終的な FC バッファーには、pH 7.5 で 500 mM トリス塩酸塩、500 mM KCl、2 mM MgCl 2、および 2 mM CaCl2 が含まれています。 - チューブFCにラベルを付け、4°Cで保管してください。
- 85 mLのRO水、10 mLの溶液T、3.73 gのKCl、および0.041 gのMgCl2 を100 mLバッファーボトルに加えます。少量の濃縮KOHを加えてpHを7.5に変更します。水を加えて最終容量を100mLにし、pHを確認します。
- 一般アクチンバッファー(GAB)を調製します。
- 485 μLのTCバッファー、10 μLの10 mM ATP、および5 μLの50 mM DTTをマイクロ遠心チューブに混合します。
注:最終的なバッファー条件は、5 mM トリス塩酸、0.2 mM CaCl 2、0.5 mM DTT、および0.2 mM ATPです。 - GABとラベル付けし、4°Cで保存してください。
- 485 μLのTCバッファー、10 μLの10 mM ATP、および5 μLの50 mM DTTをマイクロ遠心チューブに混合します。
- アクチン重合バッファー(APB)を調製します。
- 455 μLのFCバッファー、25 μLの100 mM ATP、および20 μLの50 mM DTTをマイクロ遠心チューブに混合します。
注:最終的なバッファー条件は、50 mM トリス塩酸塩、500 mM KCl、2 mM MgCl 2、2 mM CaCl 22 mM DTT、および5 mM ATPです。 - チューブにAPBのラベルを付け、4°Cで保管します。
- 455 μLのFCバッファー、25 μLの100 mM ATP、および20 μLの50 mM DTTをマイクロ遠心チューブに混合します。
- アクチンの再構成
- 凍結乾燥アクチンの1 mgバイアルに100 μLの脱イオン水を加えて、ウサギ骨格筋アクチンを再構成します。上下に穏やかにピペッティングしてよく混ぜます。5 μLのサンプルに分注し、スナップフリーズし、10 mg/mLのアクチンアリコートを-80°Cで保存します。
- ビオチン化ウサギ骨格筋アクチンを20μLのRO水を加えて再構成します。5 μLのサンプルに分注し、スナップフリーズして、1 mg/mLのビオチン化アクチンアリコートを-80°Cで保存します。
- ローダミンファロイジン安定化による非標識アクチン重合
- 10 mg/mLアクチンのバイアル1本を解凍し、氷上に置いておきます。
- 新鮮なGABバッファーを調製し、アクチンアリコートに100 μLのGABを加え、穏やかに上下にピペッティングして混合します。溶液を氷上で1時間インキュベートする。
- インキュベーション中に新鮮なAPBを準備します。インキュベーション後、アクチン溶液に11 μLのAPBを加えてアクチンをフィラメントに重合します。上下に穏やかにピペッティングしてよく混ぜます。氷上に20分間置きます。
- 5 μLのローダミン標識ファロイジンを新たに重合したアクチンフィラメント溶液に加えます。暗闇の中で氷の上に1時間放置します。
- ローダミンアクチンバイアルをアルミホイルで包んで暗所で4°Cで保管します。
注意: これらのフィラメントは最大1週間使用することをお勧めします。AFのみを含むフローセルのクイックイメージングと、一貫したフィラメントの日々の表示により、AF品質を毎日確認できます。
- Alexa Fluor 488ファロイジン安定化によるビオチン化アクチン重合
- 10 mg/mLアクチンのバイアル1本とビオチン化アクチン1本を解凍し、氷上に保ちます。
- 新鮮なGABバッファーを作成します。
- 2つのバイアルを10:1のアクチン:ビオチン化アクチン比で組み合わせます(ステップ2.8.1)。アクチン混合物に100 μLのGABを加え、上下に穏やかにピペッティングしてよく混ぜます。氷上で1時間インキュベートします。
- インキュベーション中に新鮮なAPBを作ります。
- インキュベーションステップの後、アクチン溶液に11 μLのAPBを加えてアクチンを重合します。穏やかに上下にピペッティングしてよく混ぜます。氷上で20分間インキュベートします。
- 5 μLのAlexa Fluor 488標識ファロイジンを加え、氷上で暗所で1時間インキュベートします。
- アルミホイルで包んだビオチン化アクチンバイアルを4°Cの暗所で保存します。
注:これらのフィラメントは最大1週間使用できます。
3.ミオシンとビーズの準備
- ミオシンIIの再構成
- 凍結乾燥された骨格ミオシンIIを短時間スピンダウン(~5秒)し、標準的なミニ遠心分離機を使用してチューブの底に集めます。
- RO水中で調製した1 mM DTTを100 μL添加して、ミオシンを10 mg/mLに再構成します。
- RO水中の1 mM DTTの90 μLに10 mg/mLのミオシン10 μLを加えて、ストックミオシン溶液を10倍に希釈します。少量(1〜5 μL)のアリコートを行い、スナップフリーズして、-80°Cで保存します。
注:ミオシン活性は、以前に公開された46,47のような標準的な滑走フィラメントアッセイを行うことによって確認することができる。簡単な説明については、説明を参照してください。
- ストレプトアビジンコーティングビーズの洗浄
- 20 μLの1 μmストレプトアビジンビーズを80 μLのRO水に希釈します。9,600 × g でスピンダウンし、100 μLのRO水で再構成して4回洗浄します。
- 40%振幅で2分間超音波処理し、洗浄したビーズを4°Cの回転子に保管します。
4. フローセルの準備
- 50 mLチューブに30 mLの100%エタノールを加え、200 μLの0.1%w/vポリ-l-リジンを水に加え、よく混合して、ポリ-l-リジン溶液(PLL)を調製します。
- エッチングしたカバーガラスをPLL溶液に追加し、15分間浸します。カバーガラスがチューブから引き上げられるときにカバーガラスの端にのみ触れるように注意しながら、ピンセットでカバーガラスを取り外します(図1A-Cを参照)。手袋をはめた手でカバーガラスの端をつかみます。
- エタノールが残らず、カバーガラスに残留物がなくなるまで、ろ過したエアラインでカバーガラスを乾燥させます。
- 顕微鏡スライドの中央に2枚の両面粘着テープを3〜4 mm離して貼り付けます。スライドの端からぶら下がっている余分なテープをはがすか切り取ります。
- 顕微鏡スライドの長軸に垂直なテープの上にPLLコーティングされたカバーガラス(Tを形成)を追加して、チャネルを形成します。
- 小さなチューブを使用してカバーガラスをテープに圧縮し、テープが透明になるまで顕微鏡を完全にスライドさせます(図1A)。テープに気泡がないことを確認してください。これは流路からの漏れを引き起こす可能性があるためです。
注:フローセルは10〜15μLの容量を保持できます。
5.アクトミオシンバンドル調製
- 別々のチューブで、0.5 μLの標識アクチンと300 μLのAPBを混合することにより、各タイプのアクチンフィラメント(ローダミンおよびビオチン化488標識)を600倍に希釈します。対応する標識ファロイジンをさらに5 μLずつ各チューブに加え、氷上で暗所で15分間インキュベートします。
- ビオチン化アクチン溶液に、500 mg/mL の β-D-グルコース 1 μL、25 mg/mL のグルコースオキシダーゼ 1 μL、500 単位/mL のカタラーゼ 1 μL の脱酸素システムを追加します。1 μLの100 mM ATPと1 μLの100倍希釈洗浄ストレプトアビジンビーズを加えます。ピペットチップで静かにかき混ぜます。懸濁液を4°Cのローテーターに置き、残りのアクトミオシンバンドルを組み立てます。
- 15 μLの希釈ローダミンアクチンをPLLフローセルに加えます(図1D)。余分な溶液をフローセルに通しますが、流路が乾かないようにしてください。湿度チャンバーで10分間インキュベートします。
注意: 湿度チャンバーは、底に水を加え、蓋をアルミホイルで覆って光を遮断した空のピペットチップボックスから作ることができます。 - APBで1 mg/mLのカゼイン溶液を調製します。
- 1 mg/mLのカゼインを15 μL添加して、後続の成分の非特異的結合を防ぎます(図1E)。湿度チャンバーで5分間インキュベートします。
- ステップ5.2のビオチン化アクチンおよびビーズ懸濁液に所望の濃度のミオシンを加える。ピペットチップで穏やかに攪拌し、すぐに15 μLのステップ5.2懸濁液+目的のミオシン濃度をフローセルに追加します(図1F、G)。20分間インキュベートします。フローセルの開放端をマニキュアで密封し、イメージングおよび光学トラップ実験中の蒸発を防ぎます。
注:ミオシン溶液濃度が1 μMの場合、堅牢なバンドルが得られ、アッセイの望ましいカスタマイズの出発点として使用できます( 図2を参照)。
6. 光トラップ(NT2ナノトラッカー2)を用いた力測定
注:以下のプロトコルはNT2システム専用ですが、このアッセイは、蛍光機能を備えたカスタムビルドのものを含む、他の光トラップシステムで使用できます。スライドの表面に焦点を合わせ、ビーズキャリブレーションを実行し、蛍光アクチンバンドルを見つけることによってデータを取得するという一般的なワークフローは同じです。NT2システムについては、補足図S1、補足図S2、補足図S3、補足図S4、補足図S5、補足図 S6、補足図S7 に、光トラップシステムとソフトウェアインタフェースの詳細が記載されています。
- コントロールボックスとレーザーの電源を入れます(補足図S1)。
- デスクトップ上の JPKナノトラッカー アイコンをクリックして、光トラップコンピュータソフトウェアを起動します。
- 中央にある Logitech ボタンをクリックして、リモコンをウェイクアップします(補足図S2)。
- オン/オフスイッチを切り替えて蛍光モジュールの電源を入れます(補足図S3)。
- 明視野イメージング用のフィルターキューブタレットを回します(補足図S4)。
- システムの準備ができたら、画面の左下隅にあるレーザーパワーボタンを使用してレーザーを50 mWまでオンにし、30分間安定させます(補足図S5)。
- ソフトウェア内の イルミネーション、カメラ、対物レンズ 、 ステージ移動 ボタンを順番にクリックして、実験中に表示および操作できるようにこれらのウィンドウを表示します。オン /オフ ボタンをクリックして顕微鏡照明をオンにし、バーをクリックして右端までドラッグして最大出力に設定します(補足図S5)。
- サンプル領域を開き、顕微鏡ステージからサンプルホルダーを取り外します。フローセルを追加し、金属サンプルホルダーで固定し、カバーガラス付きのスライドが底にあることを確認します。
- 下部対物レンズの中央に30 μLのRO水を加えます。ピペットの先端をレンズに触れさせないでください。サンプルステージを再挿入します。
注意: NT2システムはトラップ対物レンズとして水浸対物レンズを使用するため、ユーザーのセットアップのトラップ対物レンズによって液浸媒体が異なる場合があります。 - 水のビーズがカバーガラスに触れるまで、画面上のコントロール矢印またはリモコンのL2を使用して、下の対物レンズを上げます(補足図S5)。
- 画面上の矢印またはリモコンのR2を使用して、フローセルまでの距離の約半分に達するまで上部の目標を下げます。170 μLのRO水をフローセルの上部対物レンズの真下に追加します。水の表面張力が壊れてメニスカスが形成されるまで、上部の対物レンズを下げます。
- リモコンの矢印パッドを使用して、流路に隣接するテープの端に達するまで顕微鏡ステージを移動します。サンプル ドアを閉じます。
注意: サンプルドアを閉じたときに「カチッ」とすると、レーザーシャッターが開いたことを示します。これは、ドアが閉じている場合にのみシャッターを開くことができる安全機能です。 - 画面の「対物レンズ」ウィンドウを使用して、画面上のコントロールを使用して上矢印をクリックして、レーザー対物レンズという名前の下部の対物レンズを上に移動し、テープの端に焦点を合わせます。下の矢印をクリックして、上の目標についても同じことを行います(補足図S5)。
注:二重矢印は、目標またはステージをより速く移動します。テープの端は、カバーガラスの表面に近い大きくて見つけやすいオブジェクトであるため、焦点を合わせるために使用されます。テープ内の気泡は別のオプションです。ただし、ユーザーがサーフェスフォーカスを見つけるための自動化されたルーチンまたは好みの社内方法を持っている場合、これは必要ありません。 - テープにピントが合ったら、光学トラップの上部にあるアイリスを部分的に閉じます。虹彩のポリゴン形状が表示されるまで、上部の目標を下げます。これらのエッジに焦点を合わせ、虹彩を再度開き、 南京錠 のアイコンをクリックして対物レンズを結合します(補足図S5)。
- フローティングビーズを見つけてトラップシャッターボタンをクリックしてトラップすると、 シャッター が開き、トラップレーザーがサンプルに当たるようになります。画面上のトラップカーソルをクリックしてドラッグし、 トラップ レーザーの位置を移動します。トラップしたら、ビードを校正して、電圧測定値を力と変位に関連付けます。
- キャリブレーションボタンをクリックします。パワースペクトル解析に基づいてキャリブレーションルーチンを調整し、ソフトウェア内のコーナー周波数をX、Y、およびZ方向に合わせます(補足図S6)。
- [設定]をクリックします。ビーズの直径(1,000 nm)を入力し、ソフトウェアウィンドウの左下にあるステージの温度を入力します。(補足図S6を参照)。
- トラップ1をクリックします。X信号をクリックします。[実行]をクリックして、コーナー周波数フィットを実行します。ウィンドウ内をクリックしてドラッグし、機能のフィットを最適化します。[感度と剛性の値に使用する]をクリックします。[値の受け入れ] をクリックします。Y信号とZ信号についても繰り返します。ウィンドウを閉じます。(補足図S6を参照)。
注:等分割法や抗力法など、ユーザーによって堅牢にテストされた他の光学トラップシステムまたはカスタムビルドシステムのビードキャリブレーションルーチンも許容されます57,58。 - カバーガラスの表面でAFに結合したビーズを検索して、アクトミオシンバンドルを見つけます。
- 他の浮遊ビーズで密集していないビーズが検出された場合、蛍光イメージングによって周囲のAFを観察し、バンドルの存在を確認します。
- 共局在化された両方の蛍光AFを探して、バンドルが存在することを確認します。白色光源をオンにし、適切なフィルターキューブを使用して、タレットを回して各アクチンフィラメントをイメージングします(Alexa Fluor 488およびローダミン励起用のそれぞれ488 nmおよび532 nm励起フィルターキューブ)。 補足図S4を参照してください。
注:単一AFの蛍光強度を検証する対照実験は、単一の488および単一のローダミン標識フィラメントで構成されるバンドル、またはユーザーが使用することを選択した蛍光色素のセットに適用できるバンドルを特定するのに役立ちます。 - 確認したら、トラップシャッターボタンをクリックして、バンドルの上部フィラメントに取り付けられたビードを トラップ します。
- 画面上のコントロールを使用して、 オシロスコープ ボタンをクリックしてデータを記録します(補足図S7)。データを記録せずに測定値を視覚化するには、[ 開始]をクリックします。すべてのデータを保存するには、[ 自動保存]をクリックします。測定値を記録するには、[ 記録の開始]をクリックします。ドロップダウンメニューのX信号またはY信号から選択して、リアルタイムで視覚化するデータ(位置、力、X方向、Y方向)を選択します。xdirection は画面上で左から右、y 方向は上下であることに注意してください。 補足図S7を参照してください。
注:データは.outファイルとして保存され、各方向の時間、電圧、変位、および力が含まれます。これらのファイルは、視覚化と分析のために他のソフトウェアにエクスポートできます。
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
アクトミオシンバンドルシステムを含むフローセルは、顕微鏡スライドと両面粘着テープで作られたチャネルで区切られたエッチングされたカバーガラスで構成される標準設計です(図1)。次に、アッセイは、プロトコルに記載されている段階的な導入を使用してカバースリップから構築されます。最終的なアッセイは、テンプレートローダミン標識アクチンフィラメントで構成されています。所望のミオシン濃度( 図2 および 図3の代表的な結果には1μMを使用した)。ビオチン化、Alexa Fluor 488標識アクチンフィラメント;1μmのストレプトアビジンビーズ;酸素消去システム;ATP;および APB バッファを使用します。フローセルごとに複数のバンドルが形成され、上記のアクチン濃度はバンドル間に適切な間隔を与え、不要な相互作用がないことを保証します。これにより、フローセルごとに複数の力測定値を取得しやすくなり、データ収集効率が向上します。力プロファイルは、フローセル内およびフローセル間で再現可能である必要があります。
上記のプロトコルは市販の光学トラップセットアップの使用を対象としていますが、ここで紹介するフローセルとアッセイは、顕微鏡または顕微鏡ステージと組み合わせ、蛍光イメージング機能を備えた別の商用機器またはカスタムビルドの光学トラップセットアップに簡単に利用できます。上記のプロトコルに従ってすべてのフローセルの追加が完了すると、スライド上のアクトミオシンバンドル(図1)をすぐに測定できるようになります。フローセルを光学トラップ顕微鏡ステージに追加し、複数のビーズキャリブレーション測定値を取得し、バンドルフィラメントの蛍光共局在によってバンドルを同定します。束に結合されたビードがトラップされ、変位とそれに対応する力の測定が開始されます。ユーザーは、コンピューターのモニターでデータの取得をリアルタイムで観察できます。フローセルで使用されるミオシンの濃度に応じて、バンドルはすぐに大きな動きを示し始めるか、変位/力の増加を効果的に確認するのに30 s〜1分かかる場合があります。
代表的な力の痕跡を 図3A に示し、ミオシンモーターは安定した力のランプを示し、その後にプラトーが続きます。これらのタイプのトレースは、2〜5分かけて発生するのが一般的です。ただし、正味の力を発生しないアクトミオシンバンドルを測定することも可能です(図3B)。これらのトレースはベースラインノイズとして現れるか、90秒を超える力の実質的な正味の増加を示しません。これは、モーターの局所濃度が低く、生産的なスライドが許可されていないか、バンドルがフィラメントのプラス端とマイナス端が整列している不利な平行方向にあることが原因である可能性があります。
フローセルの内容物は、入射照明および捕捉レーザーによる劣化、時間の経過に伴うスライド上の局所的な加熱、およびラジカル酸素種の生成の影響を受けやすい可能性があるため、同じフローセルを1時間以上使用しないことを強くお勧めします。効率を最大にするには、データの取得中に別のアッセイをインキュベートすることをお勧めします。変位/フォーストレースは、光学トラッピングソフトウェアからExcel、Matlab、Igor、またはその他のデータ管理プログラムにエクスポートして、さらにフィルタリングおよび分析することができます。このような光学トラッピングアンサンブル/バンドル実験から抽出できるデータには、さまざまなアッセイ条件下でのさまざまなタイプの力生成プロファイル(ベースライン、ランプ/プラトー)、力発生速度、最大力発生、アンサンブルの動力学、ステップサイズ、ステップまたはステップチーム間の滞留時間、およびデューティ比が含まれます。また、アッセイ条件を変更して、異なるタイプのミオシンモーターの追加、アクチン結合タンパク質の追加、またはバッファー条件の変更がこれらのアンサンブル力生成特性にどのように影響するかを比較することもできます。
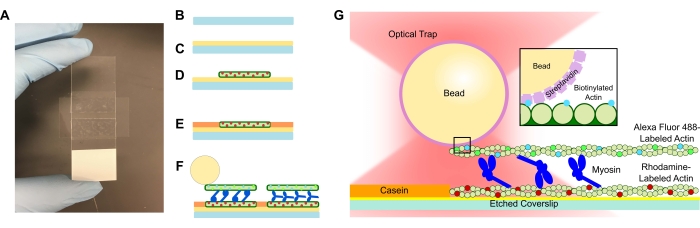
図1:アッセイ概略図。 (A-C)エッチングされたカバーガラスはポリ-L-リジンでコーティングされ、両面テープと顕微鏡スライドを使用してフローセルを形成するために使用されます。プロトコルに記載されている時限導入およびインキュベーションステップにより、ローダミン標識ファロイジン安定化アクチンが鋳型またはボトムフィラメントとして、続いて非特異的結合を防ぐためのカゼインブロッキング(E)、および(F)Alexa Fluor 488ファロイジン安定化ビオチン化アクチンが貨物または上部フィラメントとして、フィラメントが離れてスライドしてATPが導入されたときに力を発生させるミオシンIIのチーム。モーターの形状およびバンドル内の架橋の性質は、塩濃度59などのさまざまな条件下で変化する可能性があります。以前の研究では、ミオシンテールドメインがアクチンフィラメントと相互作用し、アンサンブルの運動性を遅らせる能力を持っていることが実証されています46。しかしながら、重メロミオシン実験におけるミオシンヘッドは、隣接するアクチンフィラメント60への各ヘッドの結合を実証する。(G)ストレプトアビジンビーズはトラップの光学ハンドルとして使用され、貨物ビオチン化アクチンフィラメントにのみ結合するため、スライド上に適切なバンドルが形成されていることの検証に役立ちます。この図の拡大版を表示するには、ここをクリックしてください。

図2:蛍光アクトミオシンバンドル。図1に示すバンドルアッセイ内のアクチンフィラメントおよびバンドルの4つの異なる遭遇。Alexa Fluor 488ファロイジンチャンネルを備えた上部のカーゴビオチン化アクチンフィラメントが左側に示され、ローダミンファロイジンチャネルを備えた下部のテンプレートアクチンフィラメントが右側に示されています。下部には、同じ図が目を導くのに役立つ色付きの線でオーバーレイされて示されています。(A)上部のアクチンフィラメントは、下部のアクチンフィラメントの近くに見られますが、不完全なオーバーレイがあります。これはバンドル実験には使用されません。(B)上部と下部のアクチンフィラメントは共局在しており、各フィラメントの強度は、それらがバンドル内のそれぞれ単一のフィラメントであることを確認します。これは、バンドル実験の候補として適しています。(C)自己組織化ローダミンフィラメントの大きな束が底部にあります。共局在化されている対応する上部アクチンフィラメントがある一方で、下部フィラメントが多すぎます。したがって、バンドル実験には使用されません。これは、同じタイプの複数のアクチンフィラメントが束ねられているときに、蛍光強度がどのように増加するかの例でもあります。ユーザーは、これを単一のフィラメントと同じフィラメントタイプのバンドルを判断するためのゲージとして利用できます。(D)ボトムフィラメントが存在し、対応するトップフィラメントがなく、ブリードスルーがないことも確認されています。これはバンドル実験には使用されません。Alexa Fluor 488チャンネルのフィラメントの強度が低いことに気づき、使用されているフィルターセット(Zeissのフィルターセット09)が原因であると考えています。ローダミンチャネルに使用されるフィルターセットは、ツァイスのフィルターセット43です。この図の拡大版を表示するには、ここをクリックしてください。

図3:ミオシンIIアンサンブル力の生成。 構築された in vitro アクチン構造階層内で力を発生させる骨格ミオシンIIモーターの代表的な痕跡。ミオシンモーターは、プラトーに到達して力が持続するまで(A)、またはベースライン近くで拮抗を経験するまで(B)、集合的かつ生産的に力を生成するために協力しています。 この図の拡大版を表示するには、ここをクリックしてください。
補足図S1:ブルカー/JPKナノトラッカー2光学トラップ。 (A)コンピューターモニター。(B)コンピューターのキーボード。(C)コンピュータタワー。(D)コントローラボックス。(E)レーザー電源。(F)光学トラップ光学ボックス。(G)倒立顕微鏡。(H)顕微鏡ステージへの扉。(I)明視野コントラストイメージングと微分干渉コントラストイメージングを切り替える偏光子スライダー。 このファイルをダウンロードするには、ここをクリックしてください。
補足図S2:光トラップのリモートコントロール。 (A)電動ステージを配置するためのキーパッド。(B-C)トラップ位置を調整します。(D)A、X、Bは、それぞれメインシャッター、トラップ1シャッター、トラップ2シャッターのオンとオフを切り替えます。(E)ロジクールボタンは、コントローラーをウェイクアップするために使用されます。(F)トラップ対物レンズを配置するために使用される上下ボタン。(G)検出対物レンズを配置するために使用される上下ボタン。リモコンは不要であり、これらの操作はすべてソフトウェアで実行できることに注意してください。しかし、顕微鏡のステージ環境を見ながら対物レンズやステージ位置を制御できると便利です。このファイルをダウンロードするには、ここをクリックしてください。
補足図S3:光トラップ用蛍光モジュール。 89North PhotoFluor蛍光白色光源は、倒立顕微鏡の背面に結合されています。トグルスイッチ(矢印)でオンとオフを切り替えます。 このファイルをダウンロードするには、ここをクリックしてください。
補足図S4:蛍光フィルターキューブタレット。 タレット(矢印)を回して、DIC、ローダミン、またはAlexa Fluor 488色素のイメージングに必要なフィルターキューブを使用できます。フィルターキューブを切り替えて、異なる蛍光色素を使用するためのセットアップをカスタマイズすることができます。 このファイルをダウンロードするには、ここをクリックしてください。
補足図S5:ナノトラッカー2ソフトウェア。 (A)レーザーパワーボタンとコントロール。(B)対物レンズ位置決めウィンドウ。方向矢印は、検出(上)とトラップ(下)の対物レンズを移動するために使用されます。二重矢印は、対物レンズをより高速で移動します。左下の青と赤のボタンは、目標の切り離し、元の位置に戻します。これは、顕微鏡ステージにサンプルを出し入れするときに必要です。対物レンズと南京錠のアイコンが付いた左から3番目のボタンは、対物レンズを「結合」するため、焦点が合っていてコーラー照明を達成したときに、ユーザーは捕捉対物レンズと検出対物レンズの両方をz軸で上下に移動できます。(C)顕微鏡ステージをx軸とy軸に移動するために使用されるサンプル位置決めウィンドウ。二重矢印はステージをより高速で移動します。このウィンドウは、トップメニューの上下左右矢印アイコンをクリックするとアクティブになります。(D)カメラ可視化ウィンドウ。レンチアイコンを使用して、カスタマイズされた撮影条件を設定できます。このウィンドウは、トップメニューの カメラ アイコンをクリックするとアクティブになります。(E)顕微鏡照明窓。このウィンドウは、トップメニューの 電球 アイコンをクリックするとアクティブになります。 このファイルをダウンロードするには、ここをクリックしてください。
補足図S6:キャリブレーションウィンドウ。 (ア)このウィンドウはビードキャリブレーションに使用され、トップメニューの Cal アイコンをクリックするとアクティブになります。ビードを較正するには、コーナー周波数の最適な適合がx、y、z信号で実現されます。(B)信号ごとに、左上の適切な信号ボタンを選択します。(C) 実行 をクリックし、緑色のウィンドウ(D)内をクリックしてドラッグしてフィットを最適化します。(E)フィット感に満足したら、[感度と剛性 に使用する ]をクリックします。これにより、変位をナノメートル単位で記録し、力をピコニュートン単位で記録することができます。(F)次に、左下の [値の受け入れ ]をクリックします。y 方向と z 方向についても繰り返します。 このファイルをダウンロードするには、ここをクリックしてください。
補足図S7:データ収集ウィンドウ。 このウィンドウは、位置と力のデータを取得するために使用され、ユーザーはリアルタイムで測定値を見ることができます。(A)このウィンドウは、トップメニューの x、t アイコンをクリックするとアクティブになります。(B)ユーザーは、x信号とy信号の表示を切り替えることができます。(C) [ 開始 ] をクリックして、データの視覚化を開始します。 [自動保存 ]をクリックしてデータを保存します。[記録 の開始 ]をクリックして、データの記録と保存を開始します。 このファイルをダウンロードするには、ここをクリックしてください。
Subscription Required. Please recommend JoVE to your librarian.
Discussion
アクチンフィラメントと相互作用するミオシンアンサンブルのダイナミクスを調べるために、蛍光イメージングと組み合わせた光ピンセットを使用した in vitro 研究が行われました。アクチン-ミオシン-アクチンバンドルは、筋ミオシンII、バンドルの下部とカバーガラス表面にローダミンアクチン、およびバンドルの上部に488標識されたビオチン化アクチンフィラメントを使用して組み立てられました。ウサギの筋肉由来のアクチンタンパク質を、一般的なアクチンバッファー(GAB)およびアクチン重合バッファー(APB)を使用して重合および安定化しました。GABおよびAPBは、ATP、FCバッファー、およびTCバッファーを使用して、ラボで毎日新たに調製する必要があります。筋肉ミオシンIIは、アクチン-ミオシン-アクチンサンドイッチを形成するために使用されました。ファロイジンは、アクチンフィラメントの蛍光染色、および in vitroでの安定化に使用されました。
ミオシン活性は、以前に公開されている46、47のような標準的な滑走フィラメントアッセイを行うことによって確認することができる。ミオシンIIおよびそのサブフラグメントは、様々な配向でカバーガラス表面に結合することができ、テールドメインの存在は、重いメロミオシンを使用するアッセイと比較してフィラメントの滑りを遅くすることができる46、48、49。ただし、滑空と表面の動きは引き続き観察できます。ミオシン活性のより明白なデモンストレーションは、より長いアクチンフィラメントがより小さな断片に分割され、その後複数の方向に滑空する場所で観察できる活性アクチンフィラメント破壊です。これは、表面上の高濃度の活性モーターのために起こり、複数の実験室によって観察されており、活性ミオシンモーターが存在しなければ発生しない42、50、51、52、53、54。さらに、ここで提示されるバンドルアッセイは、モーターがバンドル内で結合するようにガラス表面のカゼイン遮断を含むため、ガラスカバーガラス上の様々なモーター結合方向など、主にグライディングフィラメントアッセイに関連していた運動性の問題を軽減するのに役立ちます47、55、56。
最初のステップは、フローセル内のポリL-リジンコーティングカバーガラスに、底部またはテンプレートフィラメントとしてローダミンアクチンフィラメントを追加することです。ポリ-L-リジンは正に帯電し、アクチンは負電荷を持ち、以前の細胞骨格インビトロアッセイ調製物で使用されてきたため、ポリ-L-リジンはアクチン結合を促進するために使用されます61,62,63。バンドル形成前に、アクチン濃度を最適化するために、異なるアクチン希釈液をフローセルに添加しました。この場合、ストックから600倍が最適な希釈であり、バンドル形成に十分な数のテンプレートフィラメントが得られましたが、バンドルが個別化されるように適切な間隔がありました。希釈はAPBバッファーを用いて行った。ローダミンアクチンを添加した後、カゼインの層を加えて表面をブロックし、非特異的結合を回避しました。フローセルを30分間インキュベートし、緩衝液でインキュベートした後に洗浄して、結合していないアクチンフィラメントを洗い流した。最後に、ミオシン、488/ビオチンアクチン、およびストレプトアビジンコーティングビーズの組み合わせをフローセルに追加して、アクチン-ミオシンバンドルの形成を促進しました。ビーズ濃度は、表面結合バンドルを結合するのに十分であり、キャリブレーションを容易にするのに十分な懸濁液が存在するようなものである必要があります。ただし、ビーズ濃度が高すぎると、隣接するビーズがレーザートラップに落下して測定が中断されるため、トラップ実験中に困難が生じる可能性があります。ミオシンモーターは、スライドに注入する直前に組み合わせに追加され、ミオシンモーターが貨物または上部のビオチン化アクチンフィラメントと先制的に凝集せず、下部のローダミンを結合してビオチン化アクチンフィラメントを束ねます。
NT2光学トラップシステムは、明視野、微分干渉コントラスト(DIC)、および落射蛍光イメージングモダリティを組み合わせた商用光学トラップです。100倍/NA 1.46および63倍/NA 1.0の水浸トラップおよび検出対物レンズを備えたツァイスAxioObserver 3倒立顕微鏡と組み合わされています。このシステムは、1つのレーザートラップのクリックアンドドラッグトラップ機能を備えており、以前にリストされたモダリティのいずれかでイメージング中に使用できます。形成されたバンドルは、蛍光イメージングを用いて検出および確認される。適切なフィルターキューブ(GFP/FITCおよびTRITC/CY3)を備えた白色光源を使用すると、フィラメントイメージングを迅速に切り替えることができます。共局在AFは、光ピンセットを使用して各力測定を行う前に、異なる励起波長でAFを視覚化することによって検証されました。フィラメントは酸素消去試薬を使用しても迅速に光退色できるため、研究者はバンドル実験を実行する前に強度や露光時間などの視覚化パラメータを最適化することをお勧めします。
光学トラップを使用して力を測定し、ATPの存在下でストレプトアビジンビーズを使用してビオチン化カーゴアクチンフィラメントを結合し、力変換器としてミオシン力発生を活性化しました。光学トラップによって得られた変位および力対時間データは、分析のためにトラップソフトウェアから抽出された。しかしながら、市販のトラッピングソフトウェアは、利用することができる分析ルーチンも提供するか、または他のプログラムのカスタムアルゴリズムをユーザがプログラムして、トラッピングデータを視覚化および分析することができる。カスタム光学トラップシステムでは、ユーザーはフィルター付きの白色光源の代わりに励起レーザーを使用している場合があり、これも使用できます。また、蛍光色素は、発光スペクトルが重ならずブリードスルーを生じさせなければ、使用者が有する可能性のある既存の機器に合わせて変更することができる。
提示されたアッセイはベースラインアッセイであり、アクトミオシンアンサンブルメカニズムの領域内の研究課題に応じてユーザーがさらにカスタマイズできることに注意してください。一般的なワークフローは、有糸分裂紡錘体32、61、63、64、65、66の最小モデルを形成する微小管束アッセイなど、関心があり得る他のインビトロ細胞骨格アンサンブル系にも適用することができる。変更には、ユーザーの既存のセットアップに適したフルオロフォア標識の変更が含まれますが、これらに限定されません。ミオシン濃度、構築物、またはアイソタイプを変更する。他の側面の中でも、緩衝条件の滴定。
このアッセイを実行する際に潜在的な課題が生じる可能性があります。アクチン-ミオシンバンドルを形成する場合、アクチンバンドル内のミオシン濃度はスライド全体で均一でない場合があります。これに対応するために、スライド全体の複数のバンドルを測定して、モーター分布と力生成プロファイルが適切にサンプリングされていることを確認します。また、力データの解釈にバンドルの向きを知ることも困難です。したがって、バンドルごとに複数の試行を行う必要があります。また、光学トラップハンドルよりも小さいサイズの蛍光ゲルソリンまたはゲルソリンコーティングビーズを介してアクチンフィラメント末端標識を組み込むこともできます。蛍光イメージングを使用して、xおよびy成分の力を見て、バンドルの向きを推測することもできます。さらに、ミオシン凝集状態は、KClの急速希釈時に生じる太いフィラメントの形成を伴う緩衝液のイオン強度に大きく影響されるため、緩衝液塩濃度は適切にモニターする必要がある67,68。
グライディングアッセイなどの他のin vitro法を使用した以前の研究は、ミオシンドメインの役割を特定し、ミオシンと他のアクチン結合タンパク質との間の配置と相互作用を研究するのに役立ちました。しかしながら、これらの方法は、ミオシンを剛直な表面に結合させることが、ミオシンモーター間の協調の可能性を制限し、したがって、モーターアンサンブルが高デューティ比モードまたは低デューティ比モードにあるかどうかを決定するために生じるメカノセンシングフィードバックを制限するという欠点を有する33、35、41、69。さらに、単一ミオシンモーターネットワークを用いた光トラップでは、ミオシンモーターが互いにどのように相互作用するか、およびアクチンフィラメントとどのように相互作用するかを明確に理解することはできません。ここで開発されたプロトコルは、準拠した階層的なアクチンネットワーク内でのミオシン運動アンサンブルダイナミクスの調査を可能にします。また、濃度、アイソフォーム、バッファー環境などのモーターフィラメントアンサンブル特性の面でもカスタマイズ可能で、体系的な調査が可能です。提示されたプロトコルは、より複雑なアクトミオシンネットワークの将来の研究のためのプラットフォームであり、従来単一分子研究に使用されてきた光学トラップによって促進される変位および力発生測定の精度を維持します。
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
著者は宣言する利益相反はありません。
Acknowledgments
この研究の一部は、ミシシッピ大学大学院学生評議会研究フェローシップ(OA)、ミシシッピ大学サリーマクドネルバークスデールオナーズカレッジ(JCW、JER)、助成金番号NNX15AH78H(JCW、DNR)のミシシッピスペースグラントコンソーシアム、および助成金番号848586(DNR)のアメリカ心臓協会によってサポートされています。
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
