

Summary
Vorming van actomyosinebundels in vitro en het meten van myosine ensemblekrachtgeneratie met behulp van een optisch pincet wordt gepresenteerd en besproken.
Abstract
Myosines zijn motoreiwitten die ATP hydrolyseren om langs actinefilament (AF) tracks te stappen en zijn essentieel in cellulaire processen zoals motiliteit en spiercontractie. Om hun krachtgenererende mechanismen te begrijpen, is myosine II zowel op het niveau van één molecuul (SM) als als teams van motoren in vitro onderzocht met behulp van biofysische methoden zoals optische vangmethoden.
Deze studies toonden aan dat myosinekrachtgenererend gedrag sterk kan verschillen bij het verplaatsen van het niveau van één molecuul in een opstelling met drie kralen naar groepen motoren die samenwerken op een stijve kraal of een bedekkend oppervlak in een glijdende opstelling. Deze testconstructies laten echter niet toe om de groepsdynamiek van myosine binnen de visco-elastische structurele hiërarchie te evalueren zoals ze dat binnen een cel zouden doen. We hebben een methode ontwikkeld met behulp van optische pincetten om de mechanica van krachtgeneratie door myosine-ensembles te onderzoeken die interageren met meerdere actinefilamenten.
Deze actomyosinebundels vergemakkelijken onderzoek in een hiërarchische en conforme omgeving die motorische communicatie en ensemblekrachtoutput vastlegt. De aanpasbare aard van de test maakt het mogelijk om experimentele omstandigheden te wijzigen om te begrijpen hoe wijzigingen in het myosine-ensemble, de actinefilamentbundel of de omgeving resulteren in verschillende krachtoutputs.
Introduction
Motoreiwitten zijn essentieel voor het leven en zetten chemische energie om in mechanisch werk 1,2,3. Myosinemotoren interageren met actinefilamenten door stappen te zetten langs de filamenten die vergelijkbaar zijn met een spoor, en de dynamiek van actine-myosinenetwerken voert spiercontractie, celmotiliteit, de contractiele ring tijdens cytokinese en beweging van lading in de cel uit, naast andere essentiële taken 3,4,5,6,7,8 . Omdat myosines zoveel essentiële rollen hebben, kan falen in de functionaliteit van het myosine-actinenetwerk leiden tot ziekteontwikkeling, zoals mutaties in de myosine-zware keten die harthypercontractiliteit veroorzaken bij hypertrofische cardiomyopathie (HCM)9,10,11,12,13,14 . Bij spiercontractie werken individuele myosinemotoren met elkaar samen door als een ensemble te werken om de vereiste mechanische energie te leveren die het relatieve glijden van AFs 4,15,16,17,18 uitvoert. Myosinemotoren vormen dwarsbruggen tussen AFs en gebruiken conformatieveranderingen als gevolg van de mechanochemische cyclus om collectief naar het prikkeldraadeinde van de uitgelijnde filamenten te bewegen 17,18,19,20,21.
De ontwikkeling van kwantitatieve in vitro motiliteitstests op SM-niveau met behulp van technieken zoals optische vallen heeft het verzamelen van ongekende details over hoe individuele myosinemotoren functioneren vergemakkelijkt, inclusief het meten van SM-krachtgeneratie en stapgroottes 22,23,24,25,26,27,28,29,30 . Finer et al. ontwikkelden de "three-bead" of "dumbbell" optische trapping assay om de krachtgeneratiemechanica van enkele myosine II-motorente onderzoeken 23,31. Omdat spiermyosine II in teams werkt om AFs te contracteren, maar niet-processief is op SM-niveau, moest de optische vangtestoriëntatie worden herschikt van de klassieke motorgebonden kraalbenadering32. Om de dumbbell-assay te vormen, werden twee optische vallen gebruikt om een AF over een myosinemotor te houden die gebonden was aan een aan coverslip bevestigde kraal, en de krachtafgifte door de enkele motor werd gemeten door bewegingen van de AF binnen de val23.
SM-krachten en het gebruik van een enkele motor / single filament assay oriëntatie geven echter geen volledig beeld over het genereren van kracht op systeemniveau, omdat veel motoreiwitten, waaronder myosine II, niet geïsoleerd werken en vaak niet functioneren als een som van hun delen 15,16,17,32,33,34,35,36 . Complexere structuren die meer dan één motor omvatten die interageert met meer dan één filament zijn nodig om de synergie van de netwerken van myosine- en actinefilamenten beter te begrijpen15,32. De dumbbell assay oriëntatie is gebruikt om kleine ensemblekrachtgeneratie te onderzoeken door meerdere myosines aan een kraal te bevestigen of een myosine-dik filament aan een oppervlak te bevestigen en de motoren in staat te stellen te interageren met de gesuspendeerde AF 4,23,34,37,38,39,40.
Andere kleine ensemble-assays omvatten een in vitro filamentglijdende assay waarbij myosinemotoren op een coverslipoppervlak worden gecoat en een kraal gebonden aan een AF wordt gebruikt om de kracht te onderzoeken die wordt gegenereerd door het team van motoren 4,35,36,38,39,40,41,42,43 . In beide gevallen zijn de myosinen gebonden aan een stijf oppervlak - kraal of coverslip - en gebruiken ze één AF. In deze gevallen zijn de motoren niet in staat om vrij te bewegen of met elkaar te communiceren, noch weerspiegelt het rigide gebonden hebben van myosinen de conforme, hiërarchische omgeving waarin de motoren zouden samenwerken in het sarcomeer32. Eerdere studies hebben gesuggereerd dat myosine II zijn omgeving kan voelen en zich dienovereenkomstig kan aanpassen aan veranderende visco-elastische of motorische concentratieomstandigheden door kenmerken zoals krachtgeneratie en duty ratio 41,44,45 te veranderen. Er is dus behoefte aan het ontwikkelen van een optische vangtest die motorische communicatie en systeemcompliancy bevordert en vastlegt om een realistischer beeld te schetsen van de mechanistische onderbouwing van myosine II ensemblekrachtgeneratie.
Hier ontwikkelden we een methode om hiërarchische structuur in vitro te koppelen aan optische vangst door actomyosinebundels of sandwiches te vormen die bestaan uit meerdere myosinemotoren die interageren tussen twee actinefilamenten. Deze modulaire testgeometrie heeft de mogelijkheid om direct te onderzoeken hoe moleculaire en omgevingsfactoren de generatie van myosinekrachten beïnvloeden. Verder heeft het onderzoeken van krachtgeneratiemechanismen via deze actine-myosine-ensembles het potentieel om te helpen bij het modelleren en begrijpen hoe grootschalige cellulaire taken, zoals spiercontractie, zich voortplanten vanaf het moleculaire niveau 9,10,13.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Etsen coverslips
- Los 100 g KOH op in 300 ml 100% ethanol in een bekerglas van 1.000 ml. Roer met een roerstaaf tot het grootste deel van de KOH is opgelost.
LET OP: Geconcentreerde KOH-oplossing kan brandwonden en schade aan kleding veroorzaken. Draag handschoenen, oogbescherming en een laboratoriumjas. - Plaats coverslips afzonderlijk in coverslip reinigingsrekken.
OPMERKING: Racks zijn ontworpen met spleten die enkele coverslips uit elkaar houden om te kunnen etsen en spoelen op elk oppervlak van de coverslip, afvoergaten in de bodem en gemaakt van materiaal dat bestand is tegen de zware etsomstandigheden. Ze kunnen op maat worden gemaakt of commercieel worden gekocht. - Bereid en label drie bekers van 1.000 ml: één met 300 ml ethanol en twee bekers met 300 ml water met omgekeerde osmose (RO).
OPMERKING: Hier was RO-water afkomstig van een laboratoriumwaterzuiveraar, maar het kan ook commercieel worden gekocht als een lokale reiniger niet beschikbaar is. - Plaats elk van de vier bekers in een bad sonicator om gedurende 5 minuten te ontgassen.
- Dompel een rek met coverslips onder in het bekerglas van KOH en ethanol en soniceer gedurende 5 minuten.
- Breng het rek met coverslips over van het KOH/ethanolbeker naar het bekerglas met alleen ethanol. Dip rek op en neer in het bekerglas totdat er geen kralen zijn.
OPMERKING: Zorg ervoor dat u de coverslips niet verstoort of het rek met geweld in het bekerglas laat vallen. Dit zorgt ervoor dat de coverslips uit het rek komen of chemische spatten veroorzaken. - Breng het rek met coverslips voorzichtig over van het ethanolbeker naar een beker water en dompel op en neer totdat er geen kralen zijn.
- Dompel het rek met coverslips onder in het bekerglas met water dat nog niet is gebruikt en soniceer opnieuw gedurende 5 minuten.
- Gebruik een fles om het rek met coverslips met water te besproeien totdat het soepel van de coverslips loopt. Herhaal dit met de ethanol.
- Zet de te drogen roosters in een oven op 90 °C gedurende 20 min. Bewaar de rekken met geëtste coverslips bij kamertemperatuur in gesloten containers om besmetting voor gebruik te voorkomen.
2. Polymerisatie van actinefilamenten
- Maak oplossing T
- Voeg in een conische buis van 50 ml 3,94 g Tris-HCl en 0,147 g CaCl2 toe. Voeg RO-water toe om een totaal volume van 50 ml te maken en meng goed.
OPMERKING: De eindconcentraties van oplossing T zijn respectievelijk 500 mM Tris-HCl en 20 mM CaCl2 . - Etiketteer de buis Oplossing T en bewaar deze bij 4 °C.
- Voeg in een conische buis van 50 ml 3,94 g Tris-HCl en 0,147 g CaCl2 toe. Voeg RO-water toe om een totaal volume van 50 ml te maken en meng goed.
- Maak TC Buffer
- Meng 40 ml RO-water en 1,5 ml oplossing T in een conische buis van 50 ml. Verander de pH naar 8,0 door kleine hoeveelheden geconcentreerde KOH toe te voegen. Voeg water toe om 50 ml van de oplossing te maken en controleer de pH. Pas de pH indien nodig aan.
OPMERKING: De uiteindelijke TC-buffer bevat 5 mM Tris-HCl en 0,2 mM CaCl2 bij pH 8. - Label de buis TC en bewaar deze bij 4 °C.
- Meng 40 ml RO-water en 1,5 ml oplossing T in een conische buis van 50 ml. Verander de pH naar 8,0 door kleine hoeveelheden geconcentreerde KOH toe te voegen. Voeg water toe om 50 ml van de oplossing te maken en controleer de pH. Pas de pH indien nodig aan.
- FC Buffer maken
- Voeg 85 ml RO-water, 10 ml oplossing T, 3,73 g KCl en 0,041 g MgCl2 toe aan een bufferfles van 100 ml. Wijzig de pH naar 7,5 door kleine volumes geconcentreerd KOH toe te voegen. Voeg water toe om een eindvolume van 100 ml te maken en controleer de pH.
OPMERKING: De uiteindelijke FC-buffer bevat 500 mM Tris-HCl, 500 mM KCl, 2 mM MgCl2 en 2 mM CaCl2 bij pH 7,5. - Label de tube FC en bewaar deze bij 4 °C.
- Voeg 85 ml RO-water, 10 ml oplossing T, 3,73 g KCl en 0,041 g MgCl2 toe aan een bufferfles van 100 ml. Wijzig de pH naar 7,5 door kleine volumes geconcentreerd KOH toe te voegen. Voeg water toe om een eindvolume van 100 ml te maken en controleer de pH.
- Bereid Algemene Actin Buffer (GAB) voor.
- Meng 485 μL TC-buffer, 10 μL 10 mM ATP en 5 μL 50 mM DTT in een microcentrifugebuis.
OPMERKING: De uiteindelijke buffercondities zijn 5 mM Tris-HCl, 0,2 mM CaCl2, 0,5 mM DTT en 0,2 mM ATP. - Label het als GAB en bewaar het bij 4 °C.
- Meng 485 μL TC-buffer, 10 μL 10 mM ATP en 5 μL 50 mM DTT in een microcentrifugebuis.
- Bereid Actine Polymerisatie Buffer (APB) voor.
- Meng 455 μL FC-buffer, 25 μL 100 mM ATP en 20 μL 50 mM DTT in een microcentrifugebuis.
OPMERKING: De uiteindelijke buffercondities zijn 50 mM Tris-HCl, 500 mM KCl, 2 mM MgCl2, 2 mM CaCl2 2 mM DTT en 5 mM ATP. - Label de tube als APB en bewaar deze bij 4 °C.
- Meng 455 μL FC-buffer, 25 μL 100 mM ATP en 20 μL 50 mM DTT in een microcentrifugebuis.
- Reconstitute actine
- Reconstitueer konijnenskeletspieractine door 100 μL gedeïoniseerd water toe te voegen aan een injectieflacon van 1 mg gelyofiliseerd actine. Meng goed door voorzichtig op en neer te pipetteren. Aliquot in monsters van 5 μL, snap-freeze en bewaar de 10 mg/ml actine aliquots bij -80 °C.
- Reconstitueer gebiotinyleerd konijnenskeletspieren actine door toevoeging van 20 μL RO-water. Aliquot in monsters van 5 μL, snap-freeze en bewaar de 1 mg / ml gebiotinyleerde actine-aliquots bij -80 °C.
- Niet-gelabelde actinepolymerisatie met rhodamine phalloidine stabilisatie
- Ontdooi één injectieflacon met 10 mg/ml actine en bewaar deze op ijs.
- Bereid verse GAB-buffer, voeg 100 μL GAB toe aan de actine-aliquot en meng door voorzichtig op en neer te pipetteren. Incubeer de oplossing op ijs gedurende 1 uur.
- Bereid verse APB tijdens de incubatie. Polymeriseer na incubatie het actine in filamenten door 11 μL APB aan de actineoplossing toe te voegen. Meng goed door voorzichtig op en neer te pipetteren. Leg 20 min op ijs.
- Voeg 5 μL rhodamine-gelabelde falloïdine toe aan de vers gepolymeriseerde actinefilamentoplossing. Laat 1 uur op ijs in het donker staan.
- Bewaar de injectieflacon rhodamine actine gewikkeld in aluminiumfolie in het donker bij 4 °C.
OPMERKING: Het wordt aanbevolen om deze filamenten maximaal 1 week te gebruiken. AF-kwaliteit kan elke dag worden bevestigd door een snelle beeldvorming van een stroomcel die alleen AFs bevat en het dagelijks bekijken van consistente filamenten.
- Biotinylated actine polymerisatie met Alexa Fluor 488 phalloidin stabilisatie
- Ontdooi één injectieflacon met 10 mg/ml actine en 1 injectieflacon met 1 mg/ml gebiotinyleerd actine en bewaar ze op ijs.
- Maak een verse GAB-buffer.
- Combineer de twee injectieflacons (stap 2.8.1) in een verhouding 10:1 actine:gebiotinyleerd actine. Voeg 100 μL GAB toe aan het actinemengsel en meng goed door voorzichtig op en neer te pipetteren. Incubeer op ijs gedurende 1 uur.
- Maak verse APB tijdens de incubatie.
- Na de incubatiestap polymeriseert u het actine door 11 μL APB aan de actineoplossing toe te voegen. Meng goed door voorzichtig op en neer te pipetteren. Broed op ijs gedurende 20 minuten.
- Voeg 5 μL Alexa Fluor 488-gelabeld falloïdine toe en incubeer op ijs in het donker gedurende 1 uur.
- Bewaar de injectieflacon met biotinylated actine gewikkeld in aluminiumfolie in het donker bij 4 °C.
OPMERKING: Deze filamenten kunnen maximaal 1 week worden gebruikt.
3. Myosine en kraalbereiding
- Myosine II reconstitueren
- Kort spin naar beneden (~ 5 s) gelyofiliseerd skelet myosine II om het op de bodem van de buis te verzamelen met behulp van een standaard minicentrifuge.
- Reconstitueer de myosine tot 10 mg/ml door toevoeging van 100 μL 1 mM DTT bereid in RO-water.
- Verdun de stock myosine-oplossing 10x door 10 μL 10 mg/ml myosine toe te voegen aan 90 μL 1 mM DTT in RO-water. Maak aliquots met een klein volume (1-5 μL), vries in en bewaar bij -80 °C.
OPMERKING: Myosine-activiteit kan worden bevestigd door een standaard glijdende filamenttest uit te voeren zoals eerder gepubliceerd46,47. Zie de discussie voor een korte beschrijving.
- Reinigen van streptavidin-gecoate kralen
- Verdun 20 μL van 1 μm streptavidinkorrels tot 80 μL RO-water. Was vier keer door te spinnen bij 9.600 × g en te reconstitueren in 100 μL RO-water.
- Soniceer gedurende 2 minuten bij 40% amplitude en bewaar de gewassen kralen op een rotator bij 4 °C.
4. Stromingscelvoorbereiding
- Bereid een poly-l-lysine-oplossing (PLL) door 30 ml 100% ethanol toe te voegen aan een buis van 50 ml en 200 μL van 0,1% m / v poly-l-lysine in water toe te voegen en goed te mengen.
- Voeg een geëtste coverslip toe aan de PLL-oplossing en laat deze 15 minuten weken. Verwijder de coverslip met een pincet en zorg ervoor dat u alleen de rand van de coverslip raakt wanneer deze uit de buis wordt getrokken (zie figuur 1A-C). Pak de coverslips met een gehandschoende hand aan hun randen.
- Droog de coverslip met een gefilterde airline totdat er geen ethanol meer over is en er geen residu op de coverslip zit.
- Breng twee stukken dubbelzijdige plakband aan op het midden van een microscoopglaasje, 3-4 mm van elkaar verwijderd. Scheur of knip de overtollige tape af die aan de rand van de dia hangt.
- Voeg de PLL-gecoate coverslip toe bovenop de tape loodrecht op de lange as van de microscoopglaas (die een T vormt) om een kanaal te vormen.
- Gebruik een klein buisje om de coverslip op de tape te comprimeren en de microscoopglaas grondig te laten glijden totdat de tape transparant is (figuur 1A). Zorg ervoor dat er geen bellen in de tape zitten, omdat dit lekkage uit het stroomkanaal kan veroorzaken.
OPMERKING: De stroomcel kan een volume van 10-15 μL bevatten.
5. Actomyosine bundel voorbereiding
- Verdun in afzonderlijke buizen elk type actinefilament (rhodamine- en gebiotinyleerd 488-gelabeld) 600x door 0,5 μL van het respectieve, gelabelde actine te mengen met 300 μL APB. Voeg een extra 5 μL van het overeenkomstig gelabelde falloïdine toe aan elke buis en incubeer op ijs in het donker gedurende 15 minuten.
- Voeg aan de gebiotinyleerde actineoplossing een zuurstofopruimingssysteem toe van 1 μL bèta-D-glucose bij 500 mg/ml, 1 μL glucoseoxidase bij 25 mg/ml en 1 μL catalase bij 500 eenheden/ml. Voeg 1 μL 100 mM ATP en 1 μL 100x verdunde, gereinigde streptavidinkorrels toe. Roer voorzichtig met een pipetpunt. Zet de suspensie op een rotator op 4 °C terwijl de rest van de actomyosinebundel wordt gemonteerd.
- Voeg 15 μL van de verdunde rhodamine actine toe aan de PLL-stroomcel (figuur 1D). Leid de overtollige oplossing door de stroomcel, maar laat het stroomkanaal niet droog worden. Incubeer gedurende 10 minuten in een vochtigheidskamer.
OPMERKING: Vochtigheidskamers kunnen worden gemaakt van lege pipetpuntdozen met water toegevoegd aan de bodem en het deksel bedekt met aluminiumfolie om licht te blokkeren. - Bereid een 1 mg/ml caseïne-oplossing in APB.
- Voeg 15 μL 1 mg/ml caseïne toe om niet-specifieke binding van de volgende componenten te voorkomen (figuur 1E). Incubeer gedurende 5 minuten in een vochtigheidskamer.
- Voeg de gewenste concentratie myosine toe aan de gebiotinyleerde actine en kralensuspensie uit stap 5.2. Roer voorzichtig met de pipetpunt en voeg vervolgens onmiddellijk 15 μL van de stap 5.2 suspensie + de gewenste myosineconcentratie toe aan de stroomcel (figuur 1F,G). Incubeer gedurende 20 min. Sluit de open uiteinden van de flowcel af met nagellak om verdamping tijdens beeldvormings- en optische vangexperimenten te voorkomen.
OPMERKING: Een myosineoplossingsconcentratie van 1 μM levert robuuste bundeling op en kan worden gebruikt als uitgangspunt voor de gewenste aanpassing van de test (zie figuur 2).
6. Krachtmetingen met behulp van Optical Trap (NT2 Nanotracker2)
OPMERKING: Hoewel het onderstaande protocol specifiek voor het NT2-systeem is, kan deze test worden gebruikt met andere optische vangsystemen, inclusief systemen die op maat zijn gemaakt, die ook fluorescentiemogelijkheden hebben. De algemene workflow blijft hetzelfde als het oppervlak van de dia scherpstellen, kraalkalibraties uitvoeren en gegevens verzamelen door fluorescerende actinebundels te vinden. Voor het NT2-systeem geven Supplemental Figure S1, Supplemental Figure S2, Supplemental Figure S3, Supplemental Figure S4, Supplemental Figure S5, Supplemental Figure S6 en Supplemental Figure S7 details over het optische vangsysteem en de software-interface.
- Schakel de besturingskast en laser in (supplemental figure S1).
- Start de optische trap computersoftware door op het JPK Nanotracker-pictogram op het bureaublad te klikken.
- Activeer de afstandsbediening door op de Logitech-knop in het midden te klikken (Supplemental Figure S2).
- Schakel de fluorescentiemodule in door de aan/uit-schakelaar in te schakelen (supplemental figure S3).
- Draai de filterkubuskoepel voor brightfield-beeldvorming (aanvullende figuur S4).
- Zodra het systeem klaar is, schakelt u de laser in met behulp van de Laser Power-knop in de linkerbenedenhoek van het scherm in op 50 mW en laat u deze gedurende 30 minuten stabiliseren (aanvullende figuur S5).
- Klik achtereenvolgens op de knoppen Verlichting, Camera, Objectief en Podiumbeweging in de software om die vensters te openen voor weergave en manipulatie tijdens het experiment. Schakel de microscoopverlichting in door op de aan/uit-knop te klikken en deze in te stellen op maximaal vermogen door de balk helemaal naar rechts te klikken en te slepen (aanvullende figuur S5).
- Open het monstergebied en verwijder de monsterhouder uit het microscoopstadium. Voeg de flowcel toe, bevestig deze met de metalen monsterhouders en zorg ervoor dat de schuif met de coverslip aan de onderkant zit.
- Voeg 30 μL RO-water toe aan het midden van het onderste objectief. Laat de pipetpunt de lens niet raken. Plaats de voorbeeldfase opnieuw.
OPMERKING: Aangezien het NT2-systeem een waterdompelingsobject als vangobject gebruikt, kunnen de onderdompelingsmedia verschillen, afhankelijk van het vangdoel in de opstelling van de gebruiker. - Verhoog het onderste doel met behulp van de bedieningspijlen op het scherm of L2 op de afstandsbediening totdat de waterkraal de coverslip raakt (Aanvullende figuur S5).
- Verlaag het bovenste doel totdat ongeveer de helft van de afstand tot de stroomcel is bereikt met behulp van de pijlen op het scherm of R2 op de afstandsbediening. Voeg 170 μL RO-water toe aan de bovenkant van de stroomcel direct onder het bovenste objectief. Laat het bovenste objectief zakken totdat het de oppervlaktespanning van het water breekt en een meniscus vormt.
- Verplaats de microscooptrap met behulp van het pijlpunt op de afstandsbediening totdat de rand van de tape naast het stroomkanaal is bereikt. Sluit de monsterdeur.
OPMERKING: Een "klik" bij het sluiten van de monsterdeur geeft aan dat de lasersluiter nu open is. Dit is een veiligheidsvoorziening waarmee de sluiter alleen open kan als de deur gesloten is. - Gebruik het venster Objectief in het scherm om de rand van de tape scherp te stellen door het onderste doel met de naam Laser objective omhoog te brengen door op de bovenste pijl te klikken met behulp van de bedieningselementen op het scherm. Doe hetzelfde voor het bovenste doel door op de onderste pijl te klikken (aanvullende figuur S5).
OPMERKING: De dubbele pijlen verplaatsen het doel of de fase sneller. De rand van de tape wordt gebruikt voor het scherpstellen omdat het een groot, gemakkelijk te vinden object is dat zich dicht bij het oppervlak van de coverslip bevindt. Luchtbellen in de tape zijn een andere optie. Dit is echter niet vereist als de gebruiker een geautomatiseerde routine heeft om de oppervlaktefocus te vinden of een voorkeursmethode in huis. - Zodra de tape scherp is, sluit u de iris aan de bovenkant van de optische val gedeeltelijk. Breng het bovenste objectief naar beneden totdat de veelhoekvorm van de iris zichtbaar is. Breng die randen scherp, open de iris opnieuw en koppel vervolgens de doelstellingen door op het hangslotpictogram (aanvullende figuur S5) te klikken.
- Zoek een zwevende kraal en vang deze door op de knop Trap Shutter te klikken, waardoor de sluiter wordt geopend en de vanglaser het monster kan raken. Klik op de trapcursor op het scherm en sleep deze om de locatie van de vanglaser te verplaatsen. Eenmaal gevangen, kalibreer de kraal om spanningsmetingen te correleren met kracht en verplaatsing.
- Klik op de knop Kalibratie . Pas de kalibratieroutine aan op basis van vermogensspectra-analyse en pas de hoekfrequentie in de software in voor de X-, Y- en Z-richtingen (aanvullende figuur S6).
- Klik op Instellingen. Typ de diameter van de kraal (1.000 nm) en typ de temperatuur van het podium linksonder in het softwarevenster. (zie aanvullende figuur S6).
- Klik op Trap 1. Klik op X Signal. Klik op Uitvoeren om de hoekfrequentie te passen. Klik en sleep in het venster om de functie te optimaliseren. Klik op Use It voor gevoeligheids- en stijfheidswaarden. Klik op Waarden accepteren. Herhaal dit voor de Y- en Z-signalen. Sluit het venster. (zie aanvullende figuur S6).
OPMERKING: Kraalkalibratieroutines op andere optische vangsystemen of op maat gemaakte systemen die robuust door de gebruiker zijn getest, zoals de equipartitiemethode of sleepkrachtmethode, zijn ook acceptabel57,58. - Vind een actomyosinebundel door te zoeken naar kralen gebonden aan AFs op het oppervlak van de coverslip.
- Wanneer een kraal wordt gedetecteerd die niet wordt bevolkt door andere zwevende kralen, observeer dan de AFs eromheen door fluorescentiebeeldvorming om de aanwezigheid van een bundel te verifiëren.
- Controleer of er een bundel aanwezig is door te zoeken naar beide fluorescerende AFs die gecolokaliseerd zijn. Schakel de witte lichtbron in en gebruik de juiste filterkubus om elk actinefilament in beeld te brengen door het torentje te draaien (488 nm en 532 nm excitatiefilterkubussen voor respectievelijk Alexa Fluor 488 en rhodamine-excitatie). Zie aanvullende figuur S4.
OPMERKING: Een controle-experiment om de fluorescentie-intensiteit van afzonderlijke AFs te verifiëren, kan nuttig zijn bij het identificeren van bundels die zijn samengesteld uit enkele filamenten met een 488- en enkel rhodamine-gelabeld filament, of van toepassing op de set fluoroforen die de gebruiker kiest om te gebruiken. - Eenmaal geverifieerd, vang de kraal die aan de bovenste gloeidraad van de bundel is bevestigd door op de knop Trap Shutter te klikken.
- Gebruik de bedieningselementen op het scherm om de gegevens vast te leggen door op de knop Oscilloscoop (Aanvullende figuur S7) te klikken. Om metingen te visualiseren zonder de gegevens vast te leggen, klikt u op Start. Om alle gegevens op te slaan, klikt u op Automatisch opslaan. Om metingen vast te leggen, klikt u op Start Record. Kies welke gegevens in realtime moeten worden gevisualiseerd (positie, kracht, x-richting, y-richting) door te kiezen uit het vervolgkeuzemenu X-signaal of Y-signaal. Vergeet niet dat xdirection van links naar rechts is en dat y-direction op en neer gaat op het scherm. Zie aanvullende figuur S7.
OPMERKING: Gegevens worden opgeslagen als .out-bestanden en omvatten tijd, spanning, verplaatsing en kracht voor elke richting. Deze bestanden kunnen worden geëxporteerd naar andere software voor visualisatie en analyse.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Stromingscellen die de actomyosinebundelsystemen bevatten, hebben een standaardontwerp, bestaande uit een microscoopglaasje en een geëtste coverslip gescheiden door een kanaal gemaakt van dubbelzijdig plakband (figuur 1). De test wordt vervolgens opgebouwd uit de coverslip-up met behulp van gefaseerde introducties zoals beschreven in het protocol. De laatste test bestaat uit sjabloon rhodamine-gelabelde actinefilamenten; de gewenste myosineconcentratie (1 μM werd gebruikt voor de representatieve resultaten in figuur 2 en figuur 3); gebiotinyleerde, Alexa Fluor 488-gelabelde actinefilamenten; 1 μm streptavidin kralen; het zuurstofopruimingssysteem; ATP; en APB-buffer. Er worden meerdere bundels gevormd per stroomcel en de hierboven beschreven actineconcentraties geven voldoende afstand tussen bundels om geen ongewenste interacties te garanderen. Dit vergemakkelijkt ook het verkrijgen van meerdere krachtmetingen per stroomcel om de efficiëntie van gegevensverzameling te verhogen. Krachtprofielen moeten reproduceerbaar zijn binnen een stroomcel en van stroomcel naar stroomcel.
Hoewel het bovenstaande protocol is gericht op het gebruik van een commerciële optische vanginstallatie, kunnen de hier gepresenteerde stroomcel en assay gemakkelijk worden gebruikt voor een ander commercieel instrument of op maat gemaakte optische vangopstelling in combinatie met een microscoop- of microscoopfase en met fluorescentiebeeldvormingsmogelijkheden. Zodra alle toevoegingen van stroomcellen volgens het bovenstaande protocol zijn voltooid, zijn de actomyosinebundels op de dia (figuur 1) klaar voor onmiddellijke meting. De stroomcel wordt toegevoegd aan de optische trapmicroscoopfase, meerdere kraalkalibratiemetingen worden verkregen en bundels worden geïdentificeerd door fluorescentiecolocalisatie van de bundelfilamenten. Een kraal gebonden aan een bundel wordt gevangen en de verplaatsing en bijbehorende krachtmeting begint. De gebruiker kan de verwerving van gegevens in realtime op de computermonitor observeren. Afhankelijk van de concentratie myosine die in de stroomcel wordt gebruikt, kan de bundel onmiddellijk aanzienlijke bewegingen vertonen, of het kan 30 s-1 minuten duren om effectief een toename van verplaatsing / kracht te zien.
Een representatief krachtspoor is weergegeven in figuur 3A , waar de myosinemotoren een gestage opmars in kracht vertonen, gevolgd door een plateau. Het is typisch om dit soort sporen zich gedurende 2-5 minuten te zien ontwikkelen. Het is echter ook mogelijk om actomyosinebundels te meten die geen nettokracht genereren (figuur 3B). Deze sporen verschijnen als basisgeluid of vertonen geen substantiële netto toename van de kracht boven de 90 s. Dit is waarschijnlijk te wijten aan een lage lokale concentratie van de motor die geen productieve glijding mogelijk maakt, of de bundel bevindt zich in een ongunstige parallelle oriëntatie waarbij de plus- en minpunten van de filamenten zijn uitgelijnd.
Omdat de inhoud van de stroomcel gevoelig kan zijn voor degradatie door de invallende verlichting en vanglaser, lokale verwarming op de glijbaan in de loop van de tijd en het genereren van radicale zuurstofsoorten, wordt het ten zeerste aanbevolen om dezelfde stroomcel niet langer dan 1 uur te gebruiken. Voor maximale efficiëntie wordt voorgesteld om een andere test te laten broeden tijdens het verkrijgen van gegevens. Verplaatsings-/krachttracering kan worden geëxporteerd van de optische overvulsoftware naar Excel, Matlab, Igor of andere gegevensbeheerprogramma's voor verdere filtering en analyse. Gegevens die kunnen worden geëxtraheerd uit dergelijke optische vang ensemble / bundelexperimenten omvatten verschillende soorten krachtgeneratieprofielen (basislijn, helling / plateau) onder verschillende testomstandigheden, snelheid van krachtgeneratie, maximale krachtgeneratie, ensemblekinetisch en stapgedrag door stapgroottes en verblijftijden tussen stappen of teams van stappen, evenals de duty ratio. De gebruiker kan ook de testomstandigheden wijzigen om te vergelijken hoe het toevoegen van verschillende soorten myosinemotoren, het toevoegen van actinebindende eiwitten of het veranderen van bufferomstandigheden deze kenmerken van ensemblekrachtgeneratie beïnvloedt.
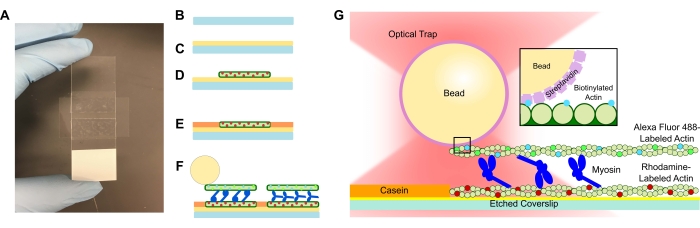
Figuur 1: Assay schematic. (A-C) Geëtste coverslips worden gecoat in poly-L-lysine en gebruikt om de flowcel te vormen met behulp van dubbelzijdige tape en een microscoopglaasje. Getimede introducties en incubatiestappen beschreven in het protocol resulteren in rhodamine-gelabelde falloidine-gestabiliseerde actine als het sjabloon of onderste filament (D), gevolgd door caseïneblokkade om niet-specifieke binding (E) te voorkomen, en (F) Alexa Fluor 488 falloidine-gestabiliseerd gestabiliseerd biotinyleerd actine als de lading of bovenste gloeidraad, en teams van myosine II die de filamenten uit elkaar schuiven en kracht genereren wanneer ATP wordt geïntroduceerd. De geometrie van de motoren en de aard van de crosslinking binnen de bundel kunnen variëren onder verschillende omstandigheden, zoals zoutconcentratie59. Eerdere studies hebben aangetoond dat het myosinestaartdomein het vermogen heeft om te interageren met actinefilamenten en langzame ensemblemotiliteit46. Myosinekoppen in zware meromyosine-experimenten tonen echter binding van elk hoofd aan aangrenzende actinefilamenten60. (G) Streptavidin-kralen worden gebruikt als het optische handvat voor de val en binden uitsluitend aan het gebiotinyleerde actinefilament van de lading, wat helpt bij het valideren dat de juiste bundels op de dia worden gevormd. Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 2: Fluorescerende actomyosinebundels. Vier verschillende ontmoetingen van actinefilamenten en bundels binnen de bundeltest in figuur 1. Het bovenste gebiotinyleerde actinefilament met het Alexa Fluor 488-falloïdinekanaal wordt links weergegeven en het onderste sjabloonactinefilament met het rhodamine-falloidinekanaal bevindt zich aan de rechterkant. Onderaan wordt dezelfde figuur weergegeven met gekleurde lijnen eroverheen om het oog te helpen begeleiden. (A) Een bovenste actinefilament bevindt zich in de buurt van een onderste actinefilament, maar heeft een onvolledige overlay. Dit zou niet worden gebruikt voor bundelexperimenten. (B) Bovenste en onderste actinefilamenten zijn gecolokaliseerd en de intensiteit van elk filament bevestigt dat het elk afzonderlijke filamenten in de bundel zijn. Dit zou een goede kandidaat zijn voor bundelexperimenten. (C) Op de bodem bevindt zich een grote bundel zelfgeassembleerde rhodaminefilamenten. Hoewel er een overeenkomstig bovenste actinefilament is dat gecolokaliseerd is, zijn er te veel onderste filamenten aanwezig; het zou dus niet worden gebruikt voor bundelexperimenten. Dit is ook een voorbeeld van hoe wanneer meerdere actinefilamenten van hetzelfde type worden gebundeld, de fluorescentie-intensiteit toeneemt. De gebruiker kan dit gebruiken als een meter voor het beoordelen van enkele filamenten versus bundels van hetzelfde filamenttype. (D) Er is een onderste gloeidraad aanwezig zonder overeenkomstige bovenste gloeidraad, wat ook bevestigt dat er geen doorbloeding is. Dit zou niet worden gebruikt voor bundelexperimenten. We merken op dat de intensiteit van de filamenten in het Alexa Fluor 488-kanaal laag is en geloven dat dit te wijten is aan de filterset die wordt gebruikt (Filter Set 09 van Zeiss). De filterset die gebruikt wordt voor het rhodamine kanaal is Filter Set 43 van Zeiss. Klik hier om een grotere versie van deze figuur te bekijken.

Figuur 3: Myosine II ensemblekrachtgeneratie. Representatieve sporen van skeletmyosine II-motoren die kracht genereren binnen de geconstrueerde in vitro actine structurele hiërarchie. De myosinemotoren werken samen om collectief en productief kracht te genereren totdat een plateau is bereikt en de kracht wordt volgehouden (A) of antagonisatie in de buurt van baseline (B) ervaren. Klik hier om een grotere versie van deze figuur te bekijken.
Aanvullende figuur S1: Bruker / JPK Nanotracker2 optische val. (A) Computermonitor. (B) Toetsenbord van de computer. (C) Computertoren. (D) Controller box. (E) Laser voeding. (F) Optische trapoptiekdoos. (G) Omgekeerde microscoop. (H) Deur naar microscoop stadium. (I) Polarizer slider om te schakelen tussen brightfield en differentiële interferentie contrast beeldvorming. Klik hier om dit bestand te downloaden.
Aanvullende figuur S2: Afstandsbediening voor optische val. (A) Toetsenbord om de gemotoriseerde fase te positioneren. (B-C) Pas de positie van de overvulling aan. (D) A, X en B schakelen respectievelijk de hoofdsluiter, trap 1 sluiter en val 2 sluiter in en uit. (E) De Logitech-knop wordt gebruikt om de controller uit de sluimerstand te halen. (F) De knoppen omhoog en omlaag die worden gebruikt om het overvuldoel te positioneren. (G) De knoppen omhoog en omlaag die worden gebruikt om het detectiedoel te positioneren. Merk op dat de afstandsbediening niet vereist is en dat al deze manipulaties in de software kunnen worden uitgevoerd. Het is echter handig om de doelstellingen en de podiumpositie te kunnen controleren terwijl u in de microscoopfase-omgeving kijkt. Klik hier om dit bestand te downloaden.
Aanvullende figuur S3: Fluorescentiemodule voor optische val. De 89North PhotoFluor fluorescentie witte lichtbron is gekoppeld aan de achterkant van de omgekeerde microscoop. Het wordt in- en uitgeschakeld met een tuimelschakelaar (pijl). Klik hier om dit bestand te downloaden.
Aanvullende figuur S4: Fluorescentiefilterkubuskoepel. Het torentje (pijl) kan worden gedraaid om de filterkubus te gebruiken die nodig is voor beeldvorming in DIC-, rhodamine- of Alexa Fluor 488-kleurstoffen. Merk op dat filterkubussen kunnen worden uitgeschakeld om de installatie aan te passen voor het gebruik van verschillende fluoroforen. Klik hier om dit bestand te downloaden.
Aanvullende figuur S5: Nanotracker2-software. (A) Laser aan / uit-knop en bediening. (B) Objectief positioneringsvenster. Richtingspijlen worden gebruikt om de doelstellingen detectie (boven) en overvulling (onder) te verplaatsen. Dubbele pijlen verplaatsen de doelen met een hogere snelheid. De blauwe en rode knop linksonder ontkoppelt de doelstellingen en trekt ze terug naar hun oorspronkelijke positie. Dit is nodig voor het nemen van monsters in en uit de microscoopfase. De derde knop van links met de doelstellingen en het hangslotpictogram "koppelt" de doelstellingen, zodat wanneer ze zowel scherp zijn als Kohler-verlichting bereiken, de gebruiker zowel de vang- als detectiedoelen op en neer kan bewegen in de z-as. (C) Monsterpositioneringsvenster dat wordt gebruikt om de microscooptrap in de x- en y-as te verplaatsen. Dubbele pijlen verplaatsen het werkgebied met een hogere snelheid. Dit venster wordt geactiveerd door te klikken op het pijltje omhoog/omlaag en pijl-links/rechts in het bovenste menu. (D) Camera visualisatie venster. Het moersleutelpictogram kan worden gebruikt om aangepaste beeldomstandigheden in te stellen. Dit venster wordt geactiveerd door op het camerapictogram in het bovenste menu te klikken. (E) Microscoop verlichting venster. Dit venster wordt geactiveerd door op het gloeilamppictogram in het bovenste menu te klikken. Klik hier om dit bestand te downloaden.
Aanvullende figuur S6: Kalibratievenster. A) Dit venster wordt gebruikt voor kraalkalibratie en wordt geactiveerd door op het cal-pictogram in het bovenste menu te klikken. Om een kraal te kalibreren, wordt een beste pasvorm van de hoekfrequentie bereikt in de x-, y- en z-signalen. (B) Kies voor elk signaal de juiste signaalknop linksboven. (C) Klik op uitvoeren en optimaliseer de pasvorm door te klikken en te slepen binnen het groene venster (D). (E) Zodra u tevreden bent met de pasvorm, klikt u op Use It voor gevoeligheid en stijfheid. Dit maakt het mogelijk om verplaatsing in nanometers en kracht in piconewtons te registreren. (F) Klik vervolgens linksonder op Waarden accepteren . Herhaal dit voor de aanwijzingen y en z. Klik hier om dit bestand te downloaden.
Aanvullende afbeelding S7: Venster Gegevensacquisitie. Dit venster wordt gebruikt om positie- en forcegegevens te verkrijgen en stelt de gebruiker in staat om de metingen in realtime te zien. (A) Dit venster wordt geactiveerd door op het x,t-pictogram in het bovenste menu te klikken. (B) De gebruiker kan schakelen tussen het bekijken van de x- en y-signalen. (C) Klik op Start om te beginnen met het visualiseren van gegevens. Klik op Automatisch opslaan om de gegevens op te slaan. Klik op Start Record om te beginnen met het opnemen en opslaan van gegevens. Klik hier om dit bestand te downloaden.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Een in vitro studie met behulp van een optisch pincet in combinatie met fluorescentie beeldvorming werd uitgevoerd om de dynamiek van myosine ensembles interactie met actine filamenten te onderzoeken. Actine-myosine-actinebundels werden geassembleerd met behulp van spiermyosine II, rhodamine-actine aan de onderkant van de bundel en op het coverslipoppervlak en 488-gelabelde, gebiotinyleerde actinefilamenten aan de bovenkant van de bundel. Actine-eiwit uit konijnenspier werd gepolymeriseerd en gestabiliseerd met behulp van algemene actinebuffers (GAB) en actinepolymerisatiebuffers (APB). GAB en APB moeten elke dag vers worden bereid in het laboratorium met behulp van ATP, FC-buffer en TC-buffer. Spiermyosine II werd gebruikt om de actine-myosine-actine sandwiches te vormen. Phalloidine werd gebruikt voor fluorescerende kleuring van de actinefilamenten, evenals stabilisatie in vitro.
Myosine-activiteit kan worden bevestigd door het uitvoeren van een standaard glijdende filamenttest zoals eerder gepubliceerd46,47. Myosine II en zijn subfragmenten kunnen in verschillende richtingen aan het coverslipoppervlak binden en de aanwezigheid van het staartdomein kan het glijden van filamenten vertragen in vergelijking met assays met zware meromyosine 46,48,49. Glijden en oppervlaktebewegingen kunnen echter nog steeds worden waargenomen. Een meer duidelijke demonstratie van myosine-activiteit is het breken van actieve actinefilamenten die kunnen worden waargenomen waarbij langere actinefilamenten worden gebroken in kleinere fragmenten die vervolgens in meerdere richtingen wegglijden. Dit gebeurt vanwege de hoge concentratie actieve motoren op het oppervlak, is waargenomen door meerdere laboratoria en komt niet voor zonder actieve myosinemotoren aanwezig 42,50,51,52,53,54. Verder helpt de hier gepresenteerde bundeltest bij het verlichten van motiliteitsproblemen die voornamelijk in verband zijn gebracht met de glijdende filamenttest, zoals de verscheidenheid aan motorbindingsoriëntaties op een glazen afdekplaat, omdat de bundeltest caseïneblokkering van het glasoppervlak omvat, zodat motoren binden binnen de bundel 47,55,56.
De eerste stap is om rhodamine actine filamenten toe te voegen als de bodem of sjabloon filament aan een poly-L-lysine gecoate coverslip in een flow cel. Poly-L-lysine wordt gebruikt om actinebinding te bevorderen, omdat poly-lysine positief geladen is, terwijl actine negatieve ladingen heeft en is gebruikt in eerdere cytoskeletale in vitro testpreparaten 61,62,63. Vóór bundelvorming werden verschillende actineverdunningen toegevoegd aan een stroomcel om de actineconcentratie te optimaliseren. In dit geval was 600x uit de voorraad de optimale verdunning die een voldoende aantal sjabloonfilamenten opleverde voor bundelvorming, maar met voldoende afstand zodat bundels werden geïndividualiseerd. De verdunning werd uitgevoerd met behulp van de APB-buffer. Het toevoegen van rhodamine actine werd gevolgd door een laag caseïne om het oppervlak te blokkeren en niet-specifieke binding te voorkomen. De stroomcel werd gedurende 30 minuten geïncubeerd en na incubatie gewassen met buffer om eventuele ongebonden actinefilamenten uit te spoelen. Ten slotte werd een combinatie van myosine, 488 / biotine-actine en streptavidin-gecoate kralen toegevoegd aan de stroomcel om de vorming van actine-myosinebundels te vergemakkelijken. De kraalconcentratie moet zodanig zijn dat er voldoende zijn om oppervlaktegebonden bundels te binden en voldoende in suspensie om kalibratie te vergemakkelijken. Een te hoge kraalconcentratie kan echter problemen veroorzaken tijdens vangexperimenten omdat naburige kralen in de laserval vallen en de meting verstoren. Myosinemotoren worden aan de combinatie toegevoegd vlak voordat deze in de dia wordt geïnjecteerd, zodat de myosinemotoren niet preventief aggregeren met de lading of het bovenste gebiotinyleerde actinefilament en zo de onderste rhodamine binden om gebiotinyleerde actinefilamenten te bundelen.
Het NT2 optische vangsysteem is een commerciële optische val met gecombineerde brightfield, differentiële interferentiecontrast (DIC) en epifluorescentiebeeldvormingsmodaliteiten. Het is gekoppeld aan een Zeiss AxioObserver 3 omgekeerde microscoop met 100x / NA 1.46 en 63x / NA 1.0 waterdompelingsvangst en detectiedoelstellingen. Het systeem is uitgerust met klik- en sleepvangmogelijkheden van één laserval en kan worden gebruikt tijdens het fotograferen in een van de eerder genoemde modaliteiten. De gevormde bundels worden gedetecteerd en bevestigd met behulp van fluorescentiebeeldvorming. Het hebben van een witte lichtbron met geschikte filterkubussen (GFP/FITC en TRITC/CY3) maakt het mogelijk om snel te schakelen tussen filamentbeeldvorming. Gecolokaliseerde AFs werden geverifieerd door de AFs op de verschillende excitatiegolflengten te visualiseren voordat elke krachtmeting werd uitgevoerd met behulp van een optisch pincet. Omdat de filamenten snel kunnen fotobleken, zelfs met een zuurstofopruimend reagens, wordt voorgesteld dat onderzoekers visualisatieparameters zoals intensiteit en blootstellingstijd optimaliseren voordat ze de bundelexperimenten uitvoeren.
Optische trapping werd gebruikt om de krachtmetingen uit te voeren, waarbij de streptavidin-kralen in aanwezigheid van ATP werden gebruikt om het gebiotinyleerde cargo actine-filament te binden en myosinekrachtgeneratie als krachttransducer te activeren. Verplaatsings- en kracht versus tijdsgegevens verkregen door optische trapping werden geëxtraheerd uit de trapping-software voor analyse. De commerciële overvulsoftware biedt echter ook analyseroutines die kunnen worden gebruikt, of aangepaste algoritmen in andere programma's kunnen door de gebruiker worden geprogrammeerd om overvulgegevens te visualiseren en te analyseren. Op aangepaste optische vangsystemen kan de gebruiker excitatielasers hebben in plaats van een witte lichtbron met filters, die ook acceptabel zijn om te gebruiken. Verder kunnen fluorescerende kleurstoffen worden gewijzigd om geschikt te zijn voor de bestaande apparatuur die een gebruiker kan hebben als de emissiespectra elkaar niet overlappen en bloeding veroorzaken.
We merken op dat de gepresenteerde test een basislijntest is die verder kan worden aangepast door de gebruiker, afhankelijk van hun onderzoeksvraag binnen het rijk van actomyosine ensemblemechanica. De algemene workflow kan ook worden toegepast op andere in vitro cytoskeletale ensemblesystemen die van belang kunnen zijn, zoals microtubule bundeltests die minimale modellen vormen van mitotische spindel 32,61,63,64,65,66. Wijzigingen kunnen omvatten, maar zijn niet beperkt tot, het wijzigen van de fluorofoorlabels die geschikt zijn voor de bestaande opstelling van de gebruiker; het veranderen van myosineconcentratie, construct of isotype; en het titreren van buffercondities, naast andere aspecten.
Potentiële uitdagingen zijn mogelijk bij het uitvoeren van deze test. Bij het vormen van de actine-myosinebundels is de myosineconcentratie in de actinebundels mogelijk niet homogeen over de dia. Om hieraan tegemoet te komen, worden meerdere bundels over de hele dia gemeten om ervoor te zorgen dat motorverdelings- en krachtgeneratieprofielen correct worden bemonsterd. Het is ook een uitdaging om te weten of dit nodig is voor de interpretatie van krachtgegevens. Er moeten dus meerdere proeven worden gedaan voor elke bundel. Men zou ook actine filament end labeling kunnen opnemen door middel van fluorescerende gelsolin of gelsolin-gecoate kralen van een kleiner formaat dan de optische vanggreep. Fluorescentiebeeldvorming kan ook worden gebruikt om te kijken naar x- en y-componentkrachten om bundeloriëntatie af te leiden. Aangezien de myosineaggregatietoestand sterk wordt beïnvloed door de ionische sterkte van de buffer met vorming van dikke filamenten die optreden bij snelle verdunning van KCl, moet de bufferzoutconcentratie bovendien op passende wijze worden gecontroleerd67,68.
Eerdere studies die andere in vitro methoden gebruikten, zoals glijdende testen, waren nuttig om de rol van myosinedomeinen te identificeren en de configuratie en interacties tussen myosine en andere actinebindende eiwitten te bestuderen. Deze methoden hebben echter een nadeel in die zin dat het binden van myosine op een stijf oppervlak het potentieel voor coördinatie tussen myosinemotoren zal beperken en dus feedback zal mechanosensen die optreedt om te bepalen of het motorensemble zich in een hoge of lage duty ratio-modusbevindt 33,35,41,69. Verder geeft optische vangst met single-myosine motornetwerken geen duidelijk inzicht in hoe myosinemotoren met elkaar en met actinefilamenten interageren. Het hier ontwikkelde protocol maakt het mogelijk om de dynamiek van myosinemotoren ensembles binnen een compliant, hiërarchisch actinenetwerk te onderzoeken. Het is ook aanpasbaar in termen van motor-filament ensemble kenmerken zoals concentratie, isovorm en bufferomgeving, naast andere aspecten, om systematisch onderzoek mogelijk te maken. Het gepresenteerde protocol is een platform voor toekomstige studies van complexere actomyosinenetwerken en handhaaft de precisie van verplaatsings- en krachtgeneratiemetingen die worden vergemakkelijkt door optische vangst die traditioneel wordt gebruikt voor studies met één molecuul.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
De auteurs hebben geen belangenconflicten te melden.
Acknowledgments
Dit werk wordt gedeeltelijk ondersteund door de University of Mississippi Graduate Student Council Research Fellowship (OA), University of Mississippi Sally McDonnell-Barksdale Honors College (JCW, JER), het Mississippi Space Grant Consortium onder subsidienummer NNX15AH78H (JCW, DNR) en de American Heart Association onder subsidienummer 848586 (DNR).
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
