

Summary
Dannelse av actomyosinbunter in vitro og måling av myosinensemblekraftgenerering ved hjelp av optisk pinsett presenteres og diskuteres.
Abstract
Myosiner er motoriske proteiner som hydrolyserer ATP for å gå langs aktinfilament (AF) spor og er essensielle i cellulære prosesser som motilitet og muskelkontraksjon. For å forstå deres kraftgenererende mekanismer har myosin II blitt undersøkt både på enkeltmolekylnivå (SM) og som team av motorer in vitro ved hjelp av biofysiske metoder som optisk fangst.
Disse studiene viste at myosin kraftgenererende oppførsel kan variere sterkt når man beveger seg fra enkeltmolekylnivået i et tre-perlearrangement til grupper av motorer som arbeider sammen på en stiv perle eller dekseloverflate i et glidearrangement. Disse analysekonstruksjonene tillater imidlertid ikke evaluering av gruppedynamikken til myosin i viskoelastisk strukturhierarki som de ville gjort i en celle. Vi har utviklet en metode som bruker optisk pinsett for å undersøke mekanikken i kraftgenerering av myosinensembler som interagerer med flere aktinfilamenter.
Disse actomyosinbuntene letter etterforskning i et hierarkisk og kompatibelt miljø som fanger opp motorisk kommunikasjon og ensemblekraftutgang. Analysens tilpassbare natur gjør det mulig å endre eksperimentelle forhold for å forstå hvordan modifikasjoner av myosinensemblet, aktinfilamentbunten eller omgivelsene resulterer i forskjellige kraftutganger.
Introduction
Motorproteiner er essensielle for livet, og konverterer kjemisk energi til mekanisk arbeid 1,2,3. Myosinmotorer interagerer med aktinfilamenter ved å ta skritt langs filamentene som ligner på et spor, og dynamikken i aktin-myosin-nettverk utfører muskelkontraksjon, cellemotilitet, kontraktilringen under cytokinese og bevegelse av last inne i cellen, blant andre viktige oppgaver 3,4,5,6,7,8 . Siden myosiner har så mange viktige roller, kan svikt i funksjonaliteten til myosin-aktinnettverket føre til sykdomsutvikling, for eksempel mutasjoner i myosin-tungkjeden som forårsaker hjertehyperkontraktilitet i hypertrofisk kardiomyopati (HCM)9,10,11,12,13,14 . I muskelkontraksjon samarbeider individuelle myosinmotorer med hverandre ved å jobbe som et ensemble for å gi den nødvendige mekaniske energien som utfører den relative glidningen av AFs 4,15,16,17,18. Myosinmotorer danner tverrbroer mellom AFs og bruker konformasjonsendringer på grunn av sin mekanokjemiske syklus for kollektivt å bevege seg mot den piggete enden av de justerte filamentene 17,18,19,20,21.
Utvikling av kvantitative in vitro-motilitetsanalyser på SM-nivå ved hjelp av teknikker som optisk fangst har gjort det lettere å samle enestående detaljer om hvordan individuelle myosinmotorer fungerer, inkludert måling av SM-kraftgenerering og trinnstørrelser 22,23,24,25,26,27,28,29,30 . Finer og medarbeidere utviklet den optiske fangstanalysen "three-bead" eller "dumbbell" for å undersøke kraftgenereringsmekanikken til enkeltmyosin II-motorer23,31. Ettersom muskelmyosin II jobber i team for å få AF, men ikke er prosessiv på SM-nivå, måtte den optiske fangstanalyseretningen omorganiseres fra den klassiske motorbundne perletilnærmingen32. For å danne manualanalysen ble to optiske feller brukt til å holde en AF over en myosinmotor bundet til en dekselfestet perle, og kraftutgangen fra den ene motoren ble målt gjennom bevegelser av AF i fellen23.
SM-krefter og bruk av en enkelt motor / enkelt filamentanalyseorientering gir imidlertid ikke et fullstendig bilde av kraftgenerering på systemnivå, siden mange motorproteiner, inkludert myosin II, ikke fungerer isolert og ofte ikke fungerer som en sum av delene 15,16,17,32,33,34,35,36 . Mer komplekse strukturer som inkluderer mer enn en motor som interagerer med mer enn ett filament er nødvendig for å bedre forstå synergien til myosin og aktinfilamenter nettverk15,32. Manualanalyseorienteringen har blitt utnyttet til å undersøke generering av små ensemblekrefter ved å ha flere myosiner festet til en perle eller bruke et myosintykt filament festet til en overflate og la motorene samhandle med den suspenderte AF 4,23,34,37,38,39,40.
Andre små ensembleanalyser inkluderer en in vitro filament glidende analyse hvor myosinmotorer er belagt på en dekseloverflate, og en perle bundet til en AF brukes til å undersøke kraften som genereres av motorteamet 4,35,36,38,39,40,41,42,43 . I begge disse tilfellene er myosinene bundet til en stiv overflate - perle eller deksel - og bruker en AF. I disse tilfellene er motorene ikke i stand til å bevege seg fritt eller kommunisere med hverandre, og det å ha myosiner som er stivt bundet, gjenspeiler heller ikke det kompatible, hierarkiske miljøet der motorene ville fungere sammen i sarcomere32. Tidligere studier har antydet at myosin II kan fornemme miljøet og tilpasse seg tilsvarende endrede viskoelastiske eller motoriske konsentrasjonsforhold ved å endre egenskaper som kraftgenerering og driftsforhold41,44,45. Dermed er det behov for å utvikle en optisk fangstanalyse som fremmer og fanger motorisk kommunikasjon og systemkompatibilitet for å male et mer realistisk bilde av det mekanistiske grunnlaget for myosin II ensemblekraftgenerering.
Her utviklet vi en metode for å koble hierarkisk struktur in vitro med optisk fangst ved å danne actomyosinbunter eller smørbrød bestående av flere myosinmotorer som interagerer mellom to aktinfilamenter. Denne modulære analysegeometrien har evnen til direkte å undersøke hvordan molekylære og miljømessige faktorer påvirker ensemble myosin kraftgenerering. Videre har undersøkelse av kraftgenereringsmekanismer gjennom disse aktin-myosinensemblene potensial til å hjelpe til med modellering og forståelse av hvordan storskala cellulære oppgaver, som muskelkontraksjon, forplanter seg opp fra molekylært nivå 9,10,13.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Etsning av deksler
- Løs opp 100 g KOH i 300 ml 100% etanol i et 1000 ml beger. Rør med rørstang til flertallet av KOH er oppløst.
FORSIKTIG: Konsentrert KOH-løsning kan forårsake brannskader og skade på klær. Bruk hansker, øyevern og en laboratoriefrakk. - Plasser deksler individuelt i dekselrengjøringsstativer.
MERK: Stativer er designet med spalter som holder enkle deksler fordelt fra hverandre for å tillate etsing og skylling på hver side av dekselet, dreneringshull i bunnen og laget av materiale som tåler de tøffe etseforholdene. De kan skreddersys eller kjøpes kommersielt. - Forbered og merk tre 1000 ml beger: en med 300 ml etanol og to beger med 300 ml omvendt osmose (RO) vann.
MERK: Her ble RO-vann hentet fra en laboratorievannrenser, men det kan også kjøpes kommersielt hvis en lokal renser ikke er tilgjengelig. - Plasser hver av de fire begerne i en badesonikator til degas i 5 min.
- Senk et stativ med deksler i begeret av KOH og etanol og sonikat i 5 min.
- Overfør stativet med deksler fra KOH / etanolbegeret til etanolbegeret. Dypp opp og ned i begeret til det ikke er perler.
MERK: Pass på at du ikke forstyrrer dekslene eller slipper stativet kraftig ned i begeret. Dette vil føre til at dekslene kommer ut av stativet eller forårsaker kjemisk sprut. - Overfør forsiktig stativet med deksler fra etanolbegeret til et vannbeger, dypp opp og ned til det ikke er perler.
- Senk stativet med deksler i begeret med vann som ikke har blitt brukt ennå, og sonikere igjen i 5 minutter.
- Bruk en flaske til å spraye stativet med deksler med vann til det renner jevnt av dekslene. Gjenta med etanol.
- Sett ristene til tørk i en ovn ved 90 °C i 20 minutter. Oppbevar stativene med etsede deksler ved romtemperatur i lukkede beholdere for å forhindre forurensning før bruk.
2. Actin filament polymerisasjon
- Lag løsning T
- I et 50 ml konisk rør, tilsett 3,94 g Tris-HCl og 0,147 g CaCl2. Tilsett RO-vann for å lage et totalt volum på 50 ml og bland godt.
MERK: De endelige konsentrasjonene av oppløsning T er henholdsvis 500 mM Tris-HCl og 20 mM CaCl2 . - Merk rørløsningen T og oppbevar den ved 4 °C.
- I et 50 ml konisk rør, tilsett 3,94 g Tris-HCl og 0,147 g CaCl2. Tilsett RO-vann for å lage et totalt volum på 50 ml og bland godt.
- Lag TC-buffer
- Bland 40 ml RO-vann og 1,5 ml løsning T i et konisk rør på 50 ml. Endre pH til 8,0 ved å tilsette små mengder konsentrert KOH. Tilsett vann for å lage 50 ml av løsningen, og kontroller pH-verdien. Juster pH-verdien om nødvendig.
MERK: Den endelige TC-bufferen inneholder 5 mM Tris-HCl og 0,2 mM CaCl2 ved pH 8. - Merk tuben TC og oppbevar den ved 4 °C.
- Bland 40 ml RO-vann og 1,5 ml løsning T i et konisk rør på 50 ml. Endre pH til 8,0 ved å tilsette små mengder konsentrert KOH. Tilsett vann for å lage 50 ml av løsningen, og kontroller pH-verdien. Juster pH-verdien om nødvendig.
- Lag FC buffer
- Tilsett 85 ml RO-vann, 10 ml oppløsning T, 3,73 g KCl og 0,041 g MgCl2 til en 100 ml bufferflaske. Endre pH til 7,5 ved å tilsette små volumer konsentrert KOH. Tilsett vann for å lage et endelig volum på 100 ml og verifiser pH.
MERK: Den endelige FC-bufferen inneholder 500 mM Tris-HCl, 500 mM KCl, 2 mM MgCl 2 og 2 mM CaCl2 ved pH 7,5. - Merk tuben FC og oppbevar den ved 4 °C.
- Tilsett 85 ml RO-vann, 10 ml oppløsning T, 3,73 g KCl og 0,041 g MgCl2 til en 100 ml bufferflaske. Endre pH til 7,5 ved å tilsette små volumer konsentrert KOH. Tilsett vann for å lage et endelig volum på 100 ml og verifiser pH.
- Forbered generell aktinbuffer (GAB).
- Bland 485 μL TC-buffer, 10 μL 10 mM ATP og 5 μL 50 mM DTT i et mikrosentrifugerør.
MERK: Endelige bufferbetingelser er 5 mM Tris-HCl, 0,2 mM CaCl 2, 0,5 mM DTT og0,2 mM ATP. - Merk den som GAB og oppbevar den ved 4 °C.
- Bland 485 μL TC-buffer, 10 μL 10 mM ATP og 5 μL 50 mM DTT i et mikrosentrifugerør.
- Forbered aktinpolymerisasjonsbuffer (APB).
- Bland 455 μL FC buffer, 25 μL 100 mM ATP og 20 μL 50 mM DTT i et mikrosentrifugerør.
MERK: De endelige bufferbetingelsene er 50 mM Tris-HCl, 500 mM KCl, 2 mM MgCl 2, 2 mM CaCl 22 mM DTT og 5 mM ATP. - Merk røret som APB og oppbevar det ved 4 °C.
- Bland 455 μL FC buffer, 25 μL 100 mM ATP og 20 μL 50 mM DTT i et mikrosentrifugerør.
- Rekonstituere aktin
- Rekonstituer kanin skjelettmuskelaktin ved å tilsette 100 μL avionisert vann til et 1 mg hetteglass med lyofilisert aktin. Bland godt ved å forsiktig pipettere opp og ned. Aliquot i 5 μL prøver, snap-freeze, og lagre 10 mg / ml actin aliquots ved -80 ° C.
- Rekonstituer biotinylert kaninskjelettmuskelaktin ved å tilsette 20 μL RO-vann. Aliquot i 5 μL prøver, snap-fryse, og lagre 1 mg / ml biotinylert aktin aliquots ved -80 ° C.
- Ikke-merket aktinpolymerisasjon med rhodamin phalloidin stabilisering
- Tine ett hetteglass med 10 mg/ml aktin og ha det på is.
- Tilbered fersk GAB-buffer, tilsett 100 μL GAB til aktin aliquot, og bland ved forsiktig pipettering opp og ned. Inkuber løsningen på is i 1 time.
- Tilbered fersk APB under inkubasjonen. Etter inkubering polymeriser aktinet i filamenter ved å tilsette 11 μL APB til aktinoppløsningen. Bland godt ved å forsiktig pipettere opp og ned. Plasser på isen i 20 min.
- Tilsett 5 μL rhodaminmerket phalloidin til den nypolymeriserte aktinfilamentløsningen. La stå på is i mørket i 1 time.
- Oppbevar hetteglasset med rhodamin aktin innpakket i aluminiumsfolie i mørket ved 4 °C.
MERK: Det foreslås å bruke disse filamentene i maksimalt 1 uke. AF-kvalitet kan bekreftes hver dag gjennom en rask avbildning av en flytcelle som bare inneholder AFs og ser konsistente filamenter fra dag til dag.
- Biotinylert aktinpolymerisasjon med Alexa Fluor 488 phalloidin stabilisering
- Tine ett hetteglass med 10 mg/ml aktin og 1 hetteglass med 1 mg/ml biotinylert aktin og holde dem på is.
- Lag fersk GAB-buffer.
- Kombiner de to hetteglassene (trinn 2.8.1) i forholdet 10:1 aktin:biotinylert aktin. Tilsett 100 μL GAB til aktinblandingen og bland godt ved forsiktig pipettering opp og ned. Inkuber på is i 1 time.
- Lag fersk APB under inkubasjonen.
- Etter inkubasjonstrinnet polymeriser aktinet ved å tilsette 11 μL APB til aktinoppløsningen. Bland godt ved pipettering opp og ned forsiktig. Inkuber på is i 20 min.
- Tilsett 5 μL Alexa Fluor 488-merket phalloidin og inkuber på is i mørket i 1 time.
- Oppbevar hetteglasset med biotinylert aktin innpakket i aluminiumsfolie i mørket ved 4 °C.
MERK: Disse filamentene kan brukes i maksimalt 1 uke.
3. Myosin og perle forberedelse
- Rekonstituere Myosin II
- Spinn kort ned (~ 5 s) lyofilisert skjelettmyosin II for å samle den i bunnen av røret ved hjelp av en standard minicentrifuge.
- Rekonstituer myosinet til 10 mg/ml ved å tilsette 100 μL 1 mM DTT fremstilt i RO-vann.
- Fortynn stammyosinoppløsningen 10x ved å tilsette 10 μL 10 mg/ml myosin til 90 μL 1 mM DTT i RO-vann. Lag aliquots med lite volum (1-5 μL), snap-freeze og oppbevar ved -80 °C.
MERK: Myosinaktivitet kan bekreftes ved å utføre en standard glidefilamentanalyse som publisert tidligere46,47. Se diskusjonen for en kort beskrivelse.
- Rengjøring av streptavidinbelagte perler
- Fortynn 20 μL av 1 μm streptavidinperler i 80 μL RO vann. Vask fire ganger ved å spinne ned på 9,600 × g og rekonstituere i 100 μL RO vann.
- Sonicat i 2 minutter ved 40% amplitude og oppbevar de vaskede perlene på en rotator ved 4 °C.
4. Forberedelse av flytceller
- Forbered en poly-l-lysinløsning (PLL) ved å tilsette 30 ml 100% etanol til et 50 ml rør og tilsette 200 μL 0,1% w / v poly-l-lysin i vann og bland godt.
- Tilsett en etset deksel til PLL-løsningen og la den trekke i 15 minutter. Fjern dekselet med pinsett, pass på at du bare berører kanten av dekselet når det trekkes opp fra røret (se figur 1A-C). Ta tak i dekslene ved kantene med en hansket hånd.
- Tørk dekselet med et filtrert flyselskap til det ikke er etanol igjen og ingen rester på dekselet.
- Påfør to stykker dobbeltsidig klebrig tape til midten av et mikroskop lysbilde, 3-4 mm fra hverandre. Riv eller klipp av overflødig tape som henger utenfor kanten av lysbildet.
- Legg til PLL-belagt deksel på toppen av båndet vinkelrett på den lange aksen til mikroskopglasset (danner en T) for å danne en kanal.
- Bruk et lite rør til å komprimere dekselleveren på båndet, og mikroskopet glir grundig til tapen er gjennomsiktig (figur 1A). Forsikre deg om at det ikke er bobler i båndet, da dette kan forårsake lekkasje fra strømningskanalen.
MERK: Flytcellen kan holde et volum på 10-15 μL.
5. Actomyosin bunt forberedelse
- I separate rør fortynnes hver type aktinfilament (rhodamin- og biotinylert 488-merket) 600x ved å blande 0,5 μL av det respektive, merkede aktinet med 300 μL APB. Tilsett ytterligere 5 μL av det tilsvarende merkede phalloidin til hvert rør og inkuber på is i mørket i 15 minutter.
- Til den biotinylerte aktinoppløsningen, tilsett et oksygenscavenging-system på 1 μL beta-D-glukose ved 500 mg / ml, 1 μL glukoseoksidase ved 25 mg / ml og 1 μL katalase ved 500 enheter / ml. Tilsett 1 μL 100 mM ATP og 1 μL 100x fortynnet, rengjort streptavidinperler. Rør forsiktig med en pipettespiss. Sett suspensjonen på en rotator ved 4 °C mens resten av actomyosinbunten monteres.
- Tilsett 15 μL av det fortynnede rhodaminaktinet til PLL-strømningscellen (figur 1D). Fukt overflødig løsning gjennom strømningscellen, men la ikke strømningskanalen bli tørr. Inkuber i 10 minutter i et fuktighetskammer.
MERK: Fuktighetskamre kan lages av tomme pipettespissbokser med vann tilsatt bunnen og lokket dekket av aluminiumsfolie for å blokkere lys. - Klargjør en 1 mg/ml kaseinoppløsning i APB.
- Tilsett 15 μL 1 mg/ml kasein for å forhindre uspesifikk binding av de påfølgende komponentene (figur 1E). Inkuber i 5 minutter i et fuktighetskammer.
- Tilsett ønsket konsentrasjon av myosin til biotinylert aktin og dråpesuspensjon fra trinn 5.2. Rør forsiktig med pipettespissen, og tilsett deretter umiddelbart 15 μL av trinn 5.2-suspensjonen + ønsket myosinkonsentrasjon til strømningscellen (figur 1F,G). Inkuber i 20 min. Forsegl de åpne endene av flytcellen med neglelakk for å forhindre fordampning under avbildning og optiske fangsteksperimenter.
MERK: En myosinløsningskonsentrasjon på 1 μM gir robust bunting og kan brukes som utgangspunkt for ønsket tilpasning av analysen (se figur 2).
6. Kraftmålinger ved hjelp av Optical Trap (NT2 Nanotracker2)
MERK: Selv om protokollen nedenfor er spesielt for NT2-systemet, kan denne analysen brukes med andre optiske overlappingssystemer, inkludert de som er spesialbygde, som også har fluorescensfunksjoner. Den generelle arbeidsflyten forblir den samme for å få overflaten av lysbildet i fokus, utføre perlekalibreringer og skaffe data ved å finne fluorescerende aktinbunter. For NT2-systemet gir supplerende figur S1, tilleggsfigur S2, tilleggsfigur S3, tilleggsfigur S4, tilleggsfigur S5, tilleggsfigur S6 og supplerende figur S7 detaljer om det optiske fangstsystemet og programvaregrensesnittet.
- Slå på kontrollboksen og laseren (tilleggsfigur S1).
- Start programvaren for optisk felle ved å klikke på JPK Nanotracker-ikonet på skrivebordet.
- Vekk fjernkontrollen ved å klikke på Logitech-knappen i midten (ekstrafigur S2).
- Slå på fluorescensmodulen ved å slå av / på-bryteren (tilleggsfigur S3).
- Vri filterkubetårnet for lysfeltavbildning (tilleggsfigur S4).
- Når systemet er klart, slår du på laseren med laserstrømknappen nederst til venstre på skjermen til 50 mW og lar den stabilisere seg i 30 minutter (tilleggsfigur S5).
- Klikk sekvensielt på knappene Belysning, Kamera, Objektiv og Scenebevegelse i programvaren for å få opp vinduene for visning og manipulering under eksperimentet. Slå på mikroskopbelysningen ved å klikke på På / Av-knappen og sett den til maksimal effekt ved å klikke og dra linjen helt til høyre (tilleggsfigur S5).
- Åpne prøveområdet og fjern prøveholderen fra mikroskopstadiet. Legg til flytcellen, fest den med metallprøveholderne, og sørg for at lysbildet med dekselet er på bunnen.
- Tilsett 30 μL RO-vann til midten av bunnmålet. Ikke la pipettespissen berøre linsen. Sett inn eksempelfasen på nytt.
MERK: Siden NT2-systemet bruker et mål for nedsenking av vann som overlappingsmål, kan nedsenkingsmediet være forskjellig avhengig av overlappingsmålet i brukerens oppsett. - Løft det nedre målet ved hjelp av kontrollpilene på skjermen eller L2 på fjernkontrollen til vannperlen berører dekselet (tilleggsfigur S5).
- Senk toppmålet til omtrent halvparten av avstanden til flytcellen er nådd ved hjelp av pilene på skjermen eller R2 på fjernkontrollen. Tilsett 170 μL RO-vann til toppen av strømningscellen direkte under toppmålet. Senk toppmålet til det bryter overflatespenningen i vannet og danner en menisk.
- Flytt mikroskoptrinnet ved hjelp av pilputen på fjernkontrollen til kanten av båndet ved siden av strømningskanalen er nådd. Lukk prøvedøren.
MERK: Et "klikk" når du lukker prøvedøren indikerer at laserlukkeren nå er åpen. Dette er en sikkerhetsfunksjon som bare lar lukkeren åpnes hvis døren er lukket. - Bruk Mål-vinduet på skjermen, og bring kanten av båndet i fokus ved å bringe det nederste målet kalt Laser Objective opp ved å klikke på den øvre pilen ved hjelp av kontrollene på skjermen. Gjør det samme for det øverste målet ved å klikke på nederste pil (tilleggsfigur S5).
MERK: De doble pilene flytter målet eller trinnet raskere. Kanten av båndet brukes til å fokusere fordi det er et stort, lett å finne objekt som er nær dekseloverflaten. Luftbobler i båndet er et annet alternativ. Dette er imidlertid ikke nødvendig hvis brukeren har en automatisert rutine for å finne overflatefokus eller en foretrukket intern metode. - Når tapen er i fokus, lukker du iris delvis øverst på den optiske fellen. Ta det øverste målet ned til polygonformen på iris er synlig. Ta disse kantene i fokus, åpne iris på nytt, og par deretter målene ved å klikke på hengelåsikonet (tilleggsfigur S5).
- Finn en flytende perle og fell den ved å klikke på Trap Shutter-knappen , som åpner lukkeren og lar fangstlaseren treffe prøven. Klikk på Trap-markøren på skjermen, og dra den for å flytte plasseringen av overlappingslaseren. Når den er fanget, kalibrer perlen for å korrelere spenningsmålinger for å tvinge og forskyves.
- Klikk på kalibreringsknappen . Juster kalibreringsrutinen basert på effektspektraanalyse og tilpass hjørnefrekvensen i programvaren for X-, Y- og Z-retningene (tilleggsfigur S6).
- Klikk på Innstillinger. Skriv inn diameteren på perlen (1000 nm), og skriv inn temperaturen på trinnet som finnes nederst til venstre i programvarevinduet. (se supplerende figur S6).
- Klikk på Trap 1. Klikk på X Signal. Klikk på Kjør for å utføre hjørnefrekvenstilpasningen. Klikk og dra i vinduet for å optimalisere funksjonstilpasningen. Klikk på Bruk den for følsomhets- og stivhetsverdier. Klikk på Godta verdier. Gjenta for Y- og Z-signalene. Lukk vinduet. (se supplerende figur S6).
MERK: Perlekalibreringsrutiner på andre optiske overlappingssystemer eller spesialbygde systemer som er robust testet av brukeren, for eksempel utstyrsmetoden eller dragkraftmetoden, er også akseptable57,58. - Finn en actomyosin bunt ved å søke etter perler bundet til AFs på overflaten av coverslip.
- Når en perle som ikke er overfylt av andre flytende perler, oppdages, må du observere AFene rundt den ved fluorescensavbildning for å verifisere tilstedeværelsen av et bunt.
- Kontroller at en pakke er til stede ved å se etter begge fluorescerende AFs colocalized. Slå på den hvite lyskilden og bruk riktig filterkube for å avbilde hvert aktinfilament ved å vri tårnet (henholdsvis 488 nm og 532 nm eksitasjonsfilterkuber for Alexa Fluor 488 og rhodamineksitasjon). Se utfyllende figur S4.
MERK: Et kontrolleksperiment for å verifisere fluorescensintensiteten til enkle AFer kan være nyttig for å identifisere bunter som består av en enkelt 488- og enkelt rhodaminmerket filamenter, eller som gjelder for hvilket sett med fluoroforer brukeren velger å bruke. - Når den er bekreftet, fanger du perlen som er festet til det øverste filamentet i bunten ved å klikke på Trap Shutter-knappen .
- Bruk skjermkontrollene til å registrere dataene ved å klikke på oscilloskopknappen (tilleggsfigur S7). For å visualisere målinger uten å registrere dataene, klikk på Start. For å lagre alle dataene, klikk på Autosave. For å registrere målinger, klikk på Start opptak. Velg hvilke data som skal visualiseres i sanntid (posisjon, kraft, x-retning, y-retning) ved å velge fra rullegardinmenyen X-signal eller Y-signal. Husk at xdirection er venstre mot høyre, og y-retning er opp og ned på skjermen. Se utfyllende figur S7.
MERK: Data lagres som .out-filer og inkluderer tid, spenning, forskyvning og kraft for hver retning. Disse filene kan eksporteres til annen programvare for visualisering og analyse.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Strømningsceller som inneholder actomyosinbuntsystemene er av standard design, bestående av et mikroskop lysbilde og en etset coverslip atskilt med en kanal laget av dobbeltsidig klebrig tape (figur 1). Analysen bygges deretter fra omslaget opp ved hjelp av iscenesatte introduksjoner som beskrevet i protokollen. Den endelige analysen består av mal rhodamin-merket actin filaments; ønsket myosinkonsentrasjon (1 μM ble brukt for de representative resultatene i figur 2 og figur 3); biotinylert, Alexa Fluor 488-merket aktinfilamenter; 1 μm streptavidin perler; oksygen scavenging system; ATP; og APB-buffer. Flere bunter vil bli dannet per strømningscelle, og aktinkonsentrasjonene beskrevet ovenfor gir tilstrekkelig avstand mellom buntene for å sikre ingen uønskede interaksjoner. Dette gjør det også enklere å oppnå flere kraftmålinger per strømningscelle for å øke datainnsamlingseffektiviteten. Kraftprofiler skal kunne reproduseres i en strømningscelle og fra strømningscelle til strømningscelle.
Mens protokollen ovenfor er rettet mot bruk av et kommersielt optisk fangstoppsett, kan flytcellen og analysen som presenteres her, enkelt brukes til et annet kommersielt instrument eller spesialbygd optisk fangstoppsett kombinert med et mikroskop- eller mikroskopstadium og har fluorescensavbildningsfunksjoner. Når alle strømningscelletilsetninger er fullført i henhold til protokollen ovenfor, er actomyosinbuntene på lysbildet (figur 1) klare for umiddelbar måling. Strømningscellen legges til det optiske fellemikroskopstadiet, flere dråpekalibreringsmålinger oppnås, og bunter identifiseres gjennom fluorescenskolokalisering av buntfilamenter. En perle bundet til en bunt er fanget, og forskyvningen og tilsvarende kraftmåling begynner. Brukeren kan observere oppkjøpet av data i sanntid på dataskjermen. Avhengig av konsentrasjonen av myosin som brukes i strømningscellen, kan bunten begynne å utvise betydelig bevegelse umiddelbart, eller det kan ta 30 s-1 min for effektivt å se en økning i forskyvning / kraft.
Et representativt kraftspor er vist i figur 3A hvor myosinmotorene viser en jevn rampe i kraft etterfulgt av et platå. Det er typisk å se denne typen spor utvikle seg over 2-5 min. Det er imidlertid også mulig å måle actomyosinbunter som ikke genererer noen nettokraft (figur 3B). Disse sporene fremstår som grunnlinjestøy eller viser ingen vesentlig netto økning i kraft over 90 s. Dette skyldes sannsynligvis en lav lokal konsentrasjon av motor som ikke tillater produktiv glidning, eller bunten er i en ugunstig parallell orientering der pluss- og minusendene av filamentene er justert.
Siden innholdet i strømningscellen kan være utsatt for nedbrytning fra innfallsbelysnings- og fangstlaseren, lokal oppvarming på lysbildet over tid og generering av radikale oksygenarter, anbefales det sterkt å ikke bruke samme strømningscelle i mer enn 1 time. For maksimal effektivitet foreslås det å ha en annen analyse som inkuberer mens du anskaffer data. Fortrengning/kraftsporing kan eksporteres fra programvaren for optisk overlapping til Excel, Matlab, Igor eller andre datahåndteringsprogrammer for videre filtrering og analyse. Data som kan ekstraheres fra slike optiske fangst ensemble / bunteksperimenter inkluderer forskjellige typer kraftgenereringsprofiler (baseline, rampe / platå) under varierende analyseforhold, hastighet på kraftgenerering, maksimal kraftgenerering, ensemblekinetisk og trinnadferd gjennom trinnstørrelser og oppholdstider mellom trinn eller lag av trinn, samt pliktforhold. Brukeren kan også endre analysebetingelsene for å sammenligne hvordan tilsetning av forskjellige typer myosinmotorer, tilsetning av aktinbindende proteiner eller endrede bufferbetingelser påvirker disse ensemblekraftgenereringsegenskapene.
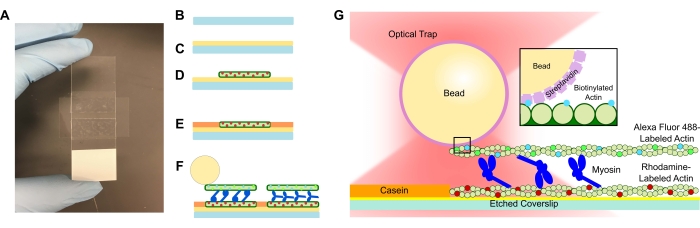
Figur 1: Analyse skjematisk. (A-C) Etsede deksler er belagt i poly-L-lysin og brukes til å danne strømningscellen ved hjelp av dobbeltsidig tape og et mikroskop lysbilde. Tidsbestemte introduksjoner og inkubasjonstrinn beskrevet i protokollen resulterer i rhodaminmerket phalloidinstabilisert aktin som mal eller bunnfilament (D), etterfulgt av kaseinblokkering for å forhindre ikke-spesifikk binding (E), og (F) Alexa Fluor 488 phalloidinstabilisert biotinylert aktin som lasten eller toppfilamentet, og lag av myosin II som skyver filamentene fra hverandre og genererer kraft når ATP introduseres. Motorenes geometri og tverrbindingens natur i bunten kan variere under forskjellige forhold, for eksempel saltkonsentrasjon59. Tidligere studier har vist at myosinhaledomenet har evnen til å interagere med aktinfilamenter og langsom ensemblemotilitet46. Imidlertid viser myosinhoder i tunge meromyosineksperimenter binding av hvert hode til tilstøtende aktinfilamenter60. (G) Streptavidin-perler brukes som det optiske håndtaket for fellen og binder seg utelukkende til lasten biotinylert aktinfilament, som hjelper til med å validere at riktige bunter dannes på lysbildet. Klikk her for å se en større versjon av dette tallet.

Figur 2: Fluorescerende actomyosinbunter. Fire ulike møter med aktinfilamenter og bunter i buntanalysen presentert i figur 1. Den øverste lasten biotinylert aktinfilament med Alexa Fluor 488 phalloidinkanalen vises til venstre, og den nederste malen actinfilament med rhodamin phalloidinkanalen er til høyre. Nederst vises den samme figuren med fargede linjer overlagt for å hjelpe øyet. (A) Et øverste aktinfilament finnes nær et bunnaktinfilament, men har et ufullstendig overlegg. Dette vil ikke bli brukt til bunteksperimenter. (B) Topp- og bunnaktinfilamenter er kolokalisert, og intensiteten til hvert filament bekrefter at de er hvert enkelt filamenter i bunten. Dette ville være en god kandidat for bunteksperimenter. (C) En stor bunt med selvmonterte rhodaminfilamenter finnes på bunnen. Mens det er et tilsvarende topp aktinfilament som er colocalized, er det for mange bunnfilamenter tilstede; Dermed ville den ikke bli brukt til bunteksperimenter. Dette er også et eksempel på hvordan når flere aktinfilamenter av samme type er buntet, øker fluorescensintensiteten. Brukeren kan bruke dette som en måler for å bedømme enkeltfilamenter versus bunter av samme filamenttype. (D) Et bunnfilament er tilstede uten tilsvarende toppfilament, og bekrefter heller ingen gjennomblødning. Dette vil ikke bli brukt til bunteksperimenter. Vi merker at intensiteten til filamentene i Alexa Fluor 488-kanalen er lav og tror det skyldes filtersettet som brukes (Filter Set 09 fra Zeiss). Filtersettet som brukes for rhodaminkanalen er filtersett 43 fra Zeiss. Klikk her for å se en større versjon av dette tallet.

Figur 3: Myosin II ensemblekraftgenerering. Representative spor av skjelettmyosin II-motorer som genererer kraft i det konstruerte in vitro aktinstrukturhierarkiet. Myosinmotorene jobber sammen for å kollektivt og produktivt generere kraft til et platå er nådd og kraften opprettholdes (A) eller opplever antagonisering nær baseline (B). Klikk her for å se en større versjon av dette tallet.
Supplerende figur S1: Bruker/JPK Nanotracker2 optisk felle. (A) Dataskjerm. (B) Tastatur på datamaskinen. (C) Datatårn. (D) Kontroller boks. (E) Laser strømforsyning. (F) Optisk felle optikk boks. (G) Omvendt mikroskop. (H) Dør til mikroskop stadium. (I) Polarisatorglidebryter for å bytte mellom brightfield og differensiell interferenskontrastbilder. Vennligst klikk her for å laste ned denne filen.
Supplerende figur S2: Fjernkontroll for optisk felle. (A) Tastatur for å plassere det motoriserte trinnet. (B-C) Juster overlappingsposisjonen. (D) A, X og B slår henholdsvis på og av hovedlukkeren, felle 1-lukkeren og felle 2-lukkeren. (E) Logitech-knappen brukes til å vekke kontrolleren. (F) Opp- og nedknappene som brukes til å plassere overlappingsmålet. (G) Opp- og nedknappene som brukes til å plassere deteksjonsmålet. Merk at fjernkontrollen ikke er nødvendig, og alle disse manipulasjonene kan oppnås i programvaren. Det er imidlertid praktisk å kunne kontrollere målene og sceneposisjonen mens du ser på mikroskopstadiet. Vennligst klikk her for å laste ned denne filen.
Supplerende figur S3: Fluorescensmodul for optisk felle. 89North PhotoFluor fluorescens hvit lyskilde er koblet til baksiden av det inverterte mikroskopet. Den slås av og på med en vippebryter (pil). Vennligst klikk her for å laste ned denne filen.
Supplerende figur S4: Fluorescensfilter kubetårn. Tårnet (pilen) kan dreies for å bruke filterkuben som er nødvendig for avbildning i DIC-, rhodamin- eller Alexa Fluor 488-fargestoffer. Merk at filterkuber kan byttes ut for å tilpasse oppsettet for bruk av forskjellige fluoroforer. Vennligst klikk her for å laste ned denne filen.
Supplerende figur S5: Nanotracker2 programvare. (A) Laserstrømknapp og kontroll. (B) Vindu for objektiv posisjonering. Retningspiler brukes til å flytte målene for gjenkjenning (øverst) og overlapping (nederst). Doble piler flytter målene med høyere hastighet. Den blå og røde knappen nederst til venstre kobler fra målene og trekker dem tilbake til sin opprinnelige posisjon. Dette er nødvendig for når du tar prøver inn og ut av mikroskopstadiet. Den tredje knappen fra venstre med målene og hengelåsikonet "parer" målene slik at når de begge er i fokus og oppnår Kohler-belysning, kan brukeren flytte både fangst- og deteksjonsmålene opp og ned i z-aksen. (C) Prøveposisjoneringsvindu som brukes til å flytte mikroskoptrinnet i x- og y-aksen. Doble piler beveger scenen med høyere hastighet. Dette vinduet aktiveres ved å klikke på pil opp/ned og venstre/høyre i toppmenyen. (D) Vindu for kameravisualisering. Skiftenøkkelikonet kan brukes til å angi tilpassede bildeforhold. Dette vinduet aktiveres ved å klikke på kameraikonet øverst i menyen. (E) Mikroskop belysning vindu. Dette vinduet aktiveres ved å klikke på lyspæreikonet i toppmenyen. Vennligst klikk her for å laste ned denne filen.
Tilleggsfigur S6: Kalibreringsvindu. (A) Dette vinduet brukes til perlekalibrering og aktiveres ved å klikke på Cal-ikonet øverst i menyen. For å kalibrere en perle, oppnås en best mulig passform av hjørnefrekvensen i x-, y- og z-signalene. (B) For hvert signal velger du riktig signalknapp øverst til venstre. (C) Klikk på kjør og optimaliser passformen ved å klikke og dra i det grønne vinduet (D). (E) Når du er fornøyd med passformen, klikker du på Bruk den for følsomhet og stivhet. Dette vil tillate opptak av forskyvning i nanometer og kraft i piconewtons. (F) Klikk deretter på Godta verdier nederst til venstre. Gjenta for y- og z-retningene. Vennligst klikk her for å laste ned denne filen.
Supplerende figur S7: Datainnsamlingsvinduet. Dette vinduet brukes til å skaffe posisjons- og kraftdata og lar brukeren se målingene i sanntid. (A) Dette vinduet aktiveres ved å klikke på x,t-ikonet i toppmenyen. (B) Brukeren kan bytte mellom å se x- og y-signalene. (C) Klikk på Start for å begynne å visualisere data. Klikk på Autosave for å lagre dataene. Klikk på Start opptak for å begynne å registrere og lagre data. Vennligst klikk her for å laste ned denne filen.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
En in vitro-studie med optisk pinsett kombinert med fluorescensavbildning ble utført for å undersøke dynamikken i myosinensembler som interagerer med aktinfilamenter. Actin-myosin-aktinbunter ble satt sammen ved hjelp av muskelmyosin II, rhodamin aktin i bunnen av bunten og på dekseloverflaten, og 488-merkede, biotinylerte aktinfilamenter på toppen av bunten. Actinprotein fra kaninmuskel ble polymerisert og stabilisert ved bruk av generelle aktinbuffere (GAB) og aktinpolymeriserende buffere (APB). GAB og APB må tilberedes hver dag i laboratoriet ved hjelp av ATP, FC-buffer og TC-buffer. Muskelmyosin II ble brukt til å danne aktin-myosin-aktinsmørbrød. Phalloidin ble brukt til fluorescerende farging av aktinfilamenter, samt stabilisering in vitro.
Myosinaktivitet kan bekreftes ved å utføre en standard glidefilamentanalyse som publisert tidligere46,47. Myosin II og dets underfragmenter kan binde seg til dekseloverflaten i en rekke retninger, og tilstedeværelsen av haledomenet kan bremse filamentglidning sammenlignet med analyser ved bruk av tung meromyosin46,48,49. Imidlertid kan gliding og overflatebevegelse fortsatt observeres. En mer tydelig demonstrasjon av myosinaktivitet er aktiv aktinfilamentbrudd som kan observeres der lengre aktinfilamenter brytes i mindre fragmenter som deretter glir bort i flere retninger. Dette skjer på grunn av den høye konsentrasjonen av aktive motorer på overflaten, har blitt observert av flere laboratorier, og forekommer ikke uten aktive myosinmotorer tilstede 42,50,51,52,53,54. Videre hjelper buntanalysen som presenteres her til å lindre motilitetsproblemer som primært har vært knyttet til glidefilamentanalysen, for eksempel forskjellige motorbindingsretninger på et glassdeksel, fordi buntanalysen innebærer kaseinblokkering av glassoverflaten slik at motorer binder seg innenfor bunten 47,55,56.
Det første trinnet er å legge til rhodamin actin filamenter som bunnen eller malfilamentet til en poly-L-lysinbelagt coverslip i en strømningscelle. Poly-L-lysin brukes til å fremme aktinbinding siden poly-lysin er positivt ladet mens aktin har negative ladninger og har blitt brukt i tidligere cytoskeletale in vitro analysepreparater61,62,63. Før buntdannelse ble forskjellige aktinfortynninger tilsatt til en strømningscelle for å optimalisere aktinkonsentrasjonen. I dette tilfellet var 600x fra lageret den optimale fortynningen som ga et tilstrekkelig antall malfilamenter for buntdannelse, men med tilstrekkelig avstand slik at bunter ble individualisert. Fortynning ble utført ved hjelp av APB-bufferen. Tilsetning av rhodamin aktin ble etterfulgt av et lag kasein for å blokkere overflaten og unngå uspesifikk binding. Strømningscellen ble inkubert i 30 minutter og vasket etter inkubering med buffer for å vaske ut eventuelle ubundne aktinfilamenter. Til slutt ble en kombinasjon av myosin, 488/biotin aktin og streptavidinbelagte perler tilsatt til strømningscellen for å lette dannelsen av aktin-myosinbunt. Dråpekonsentrasjonen skal være slik at det er nok til å binde overflatebundne bunter og nok i suspensjon for å lette kalibreringen. Imidlertid kan for høy dråpekonsentrasjon føre til vanskeligheter under fangsteksperimenter på grunn av at naboperler faller inn i laserfellen og forstyrrer målingen. Myosinmotorer tilsettes kombinasjonen rett før den injiseres i lysbildet, slik at myosinmotorene ikke preemptively aggregerer med lasten eller topp biotinylert aktinfilament og vil dermed binde bunnen rhodamin for å bunte biotinylerte aktinfilamenter.
NT2 optisk fangstsystem er en kommersiell optisk felle med kombinert brightfield, differensiell interferenskontrast (DIC) og epifluorescensavbildningsmodaliteter. Det er kombinert med et Zeiss AxioObserver 3 invertert mikroskop med 100x / NA 1.46 og 63x / NA 1.0 vann nedsenking fangst og deteksjon mål. Systemet er utstyrt med klikk- og dra-overlappingskapasitet for en laserfelle og kan brukes under avbildning i noen av de listede modalitetene tidligere. De dannede buntene oppdages og bekreftes ved bruk av fluorescensbilder. Å ha en hvit lyskilde med passende filterkuber (GFP / FITC og TRITC / CY3) muliggjør rask bytte mellom filamentbilder. Colocalized AFs ble verifisert ved å visualisere AFs på de forskjellige eksitasjonsbølgelengdene før du tar hver kraftmåling ved hjelp av optisk pinsett. Siden filamentene kan fotobleke raskt selv med et oksygenscavenging-reagens, foreslås det at forskere optimaliserer visualiseringsparametere som intensitet og eksponeringstid før de utfører bunteksperimentene.
Optisk fangst ble brukt til å ta kraftmålingene, ved hjelp av streptavidinperlene i nærvær av ATP for å binde det biotinylerte lastaktinfilamentet og aktivere myosinkraftgenerering som krafttransduser. Forskyvnings- og kraft-mot-tid-data innhentet ved optisk overlapping ble hentet fra overlappingsprogramvaren for analyse. Den kommersielle overlappingsprogramvaren inneholder imidlertid også analyserutiner som kan brukes, eller egendefinerte algoritmer i andre programmer kan programmeres av brukeren til å visualisere og analysere overlappingsdata. På tilpassede optiske fangstsystemer kan brukeren ha eksitasjonslasere i stedet for en hvit lyskilde med filtre, som også er akseptable å bruke. Videre kan fluorescerende fargestoffer endres for å være tilpasset det eksisterende utstyret en bruker kan ha hvis utslippsspektrene ikke overlapper og forårsaker gjennomblødning.
Vi bemerker at analysen som presenteres er en grunnlinjeanalyse som kan tilpasses ytterligere av brukeren, avhengig av deres forskningsspørsmål innen actomyosin ensemblemekanikk. Den generelle arbeidsflyten kan også brukes på andre in vitro cytoskeletale ensemblesystemer som kan være av interesse, for eksempel mikrotubulibuntanalyser som danner minimale modeller av mitotisk spindel 32,61,63,64,65,66. Modifikasjoner kan inkludere, men er ikke begrenset til, endring av fluoroforetikettene som passer til brukerens eksisterende oppsett; endre myosinkonsentrasjon, konstruksjon eller isotype; og titrering av bufferbetingelser, blant annet.
Potensielle utfordringer er mulige når du utfører denne analysen. Ved dannelse av aktin-myosinbuntene kan det hende at myosinkonsentrasjonen i aktinbuntene ikke er homogen over lysbildet. For å imøtekomme dette vil flere bunter over hele lysbildet måles for å sikre at motorfordeling og kraftgenereringsprofiler blir riktig samplet. Det er også utfordrende å vite buntorientering hvis dette er nødvendig for tolkning av kraftdata. Dermed bør det tas flere forsøk for hver pakke. Man kan også inkorporere actinfilamentendemerking gjennom fluorescerende gelsolin eller gelsolinbelagte perler av mindre størrelse enn det optiske fangsthåndtaket. Fluorescensavbildning kan også brukes til å se på x- og y-komponentkrefter for å utlede buntorientering. Videre, ettersom myosinaggregeringstilstanden er sterkt påvirket av bufferens ionstyrke med dannelse av tykke filamenter som oppstår ved rask fortynning av KCl, bør buffersaltkonsentrasjonen overvåkes hensiktsmessig67,68.
Tidligere studier som brukte andre in vitro-metoder som glidende analyser var nyttige for å identifisere rollen som myosindomener og studere konfigurasjonen og interaksjonene mellom myosin og andre aktinbindende proteiner. Imidlertid har disse metodene en ulempe ved at binding av myosin på en stiv overflate vil begrense potensialet for koordinering mellom myosinmotorer og dermed mekanosenserende tilbakemelding som oppstår for å avgjøre om motorensemblet er i høy eller lav driftsforholdsmodus 33,35,41,69. Videre gir optisk fangst med enkelt-myosinmotoriske nettverk ikke en klar forståelse av hvordan myosinmotorer interagerer med hverandre og med aktinfilamenter. Protokollen som er utviklet her, gjør det mulig å undersøke myosinmotorisk ensembledynamikk innenfor et kompatibelt, hierarkisk aktinnettverk. Det kan også tilpasses når det gjelder motorfilamentensembleegenskaper som konsentrasjon, isoform og buffermiljø, blant annet for å muliggjøre systematisk undersøkelse. Den presenterte protokollen er en plattform for fremtidige studier av mer komplekse actomyosin-nettverk og opprettholder presisjonen av forskyvnings- og kraftgenereringsmålinger tilrettelagt av optisk fangst som tradisjonelt har blitt brukt til enkeltmolekylstudier.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Forfatterne har ingen interessekonflikter å oppgi.
Acknowledgments
Dette arbeidet støttes delvis av University of Mississippi Graduate Student Council Research Fellowship (OA), University of Mississippi Sally McDonnell-Barksdale Honors College (JCW, JER), Mississippi Space Grant Consortium under tilskuddsnummer NNX15AH78H (JCW, DNR) og American Heart Association under tilskuddsnummer 848586 (DNR).
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
