

Summary
Bildning av aktomyosinbuntar in vitro och mätning av myosinensemblekraftgenerering med hjälp av optisk pincett presenteras och diskuteras.
Abstract
Myosiner är motorproteiner som hydrolyserar ATP för att gå längs aktinfilament (AF) spår och är väsentliga i cellulära processer såsom rörlighet och muskelkontraktion. För att förstå deras kraftgenererande mekanismer har myosin II undersökts både på enmolekylär (SM) nivå och som team av motorer in vitro med biofysiska metoder såsom optisk fångst.
Dessa studier visade att myosinkraftgenererande beteende kan skilja sig mycket när man rör sig från enmolekylnivån i ett tre-pärlarrangemang till grupper av motorer som arbetar tillsammans på en styv pärla eller täckglidyta i ett glidarrangemang. Dessa analyskonstruktioner tillåter emellertid inte utvärdering av gruppdynamiken hos myosin inom viskoelastisk strukturell hierarki som de skulle göra inom en cell. Vi har utvecklat en metod med optisk pincett för att undersöka mekaniken för kraftgenerering av myosinensembler som interagerar med flera aktinfilament.
Dessa aktomysinbuntar underlättar undersökning i en hierarkisk och kompatibel miljö som fångar motorisk kommunikation och ensemblekraftutgång. Analysens anpassningsbara natur gör det möjligt att ändra experimentella förhållanden för att förstå hur modifieringar av myosinensemblen, aktinfilamentbunten eller den omgivande miljön resulterar i olika kraftutgångar.
Introduction
Motorproteiner är livsnödvändiga och omvandlar kemisk energi till mekaniskt arbete 1,2,3. Myosinmotorer interagerar med aktinfilament genom att ta steg längs filamenten som liknar ett spår, och dynamiken i aktin-myosinnätverk utför muskelkontraktion, cellmotilitet, kontraktilringen under cytokinese och rörelse av last inuti cellen, bland andra väsentliga uppgifter 3,4,5,6,7,8 . Eftersom myosiner har så många viktiga roller kan misslyckande i funktionaliteten hos myosin-aktinnätverket leda till sjukdomsutveckling, såsom mutationer i myosin-tungkedjan som orsakar hjärthyperkontraktilitet vid hypertrofisk kardiomyopati (HCM)9,10,11,12,13,14 . Vid muskelkontraktion samarbetar enskilda myosinmotorer med varandra genom att arbeta som ett ensemble för att tillhandahålla den erforderliga mekaniska energin som utför den relativa glidningen av AFs 4,15,16,17,18. Myosinmotorer bildar korsbroar mellan AF och använder konformationsförändringar på grund av dess mekanokemiska cykel för att kollektivt röra sig mot den taggiga änden av de inriktade filamenten 17,18,19,20,21.
Utveckling av kvantitativa in vitro-motilitetsanalyser på SM-nivå med hjälp av tekniker som optisk fångst har underlättat insamling av oöverträffade detaljer om hur enskilda myosinmotorer fungerar, inklusive mätning av SM-kraftgenerering och stegstorlekar 22,23,24,25,26,27,28,29,30 . utvecklade den optiska fångstanalysen "tre-pärla" eller "hantel" för att undersöka kraftgenereringsmekaniken hos enstaka myosin II-motorer23,31. Eftersom muskelmyosin II fungerar i team för att kontraktera AF men inte är processiv på SM-nivå, måste den optiska fångstanalysorienteringen omarrangeras från den klassiska motorbundna pärlmetoden32. För att bilda hantelanalysen användes två optiska fällor för att hålla en AF över en myosinmotor bunden till en täckglasfäst pärla, och kraftutgången från den enda motorn mättes genom AF-rörelser inom fällan23.
SM-krafter och användning av en enda motor/ enda filamentanalysorientering ger dock inte en fullständig bild av kraftgenerering på systemnivå eftersom många motorproteiner, inklusive myosin II, inte fungerar isolerat och ofta inte fungerar som en summa av deras delar 15,16,17,32,33,34,35,36 . Mer komplexa strukturer som inkluderar mer än en motor som interagerar med mer än ett filament är nödvändiga för att bättre förstå synergin mellan myosin- och aktinfilamentens nätverk 15,32. Hantelanalysorienteringen har utnyttjats för att undersöka generering av små ensemblekrafter genom att ha flera myosiner fästa vid en pärla eller använda ett myosintjockt filament fäst vid en yta och låta motorerna interagera med den upphängda AF 4,23,34,37,38,39,40.
Andra små ensembleanalyser inkluderar en in vitro-filamentglidningsanalys där myosinmotorer beläggs på en täckbladsyta, och en pärla bunden till en AF används för att undersöka kraften som genereras av motorteamet4,35,36,38,39,40,41,42,43 . I båda dessa fall är myosinerna bundna till en stel yta - pärla eller täckglas - och använder en AF. I dessa fall kan motorerna inte röra sig fritt eller kommunicera med varandra, och inte heller återspeglar myosiner styvt bundna den kompatibla, hierarkiska miljön där motorerna skulle arbeta tillsammans i sarcomere32. Tidigare studier har föreslagit att myosin II kan känna av sin miljö och anpassa sig därefter till förändrade viskoelastiska eller motoriska koncentrationsförhållanden genom att ändra egenskaper som kraftgenerering och tullförhållande41,44,45. Således finns det ett behov av att utveckla en optisk fångstanalys som främjar och fångar motorisk kommunikation och systemkompatibilitet för att måla en mer realistisk bild av de mekanistiska grunderna för myosin II-ensemblekraftgenerering.
Här utvecklade vi en metod för att koppla hierarkisk struktur in vitro med optisk fångst genom att bilda aktomyosinbuntar eller smörgåsar bestående av flera myosinmotorer som interagerar mellan två aktinfilament. Denna modulära analysgeometri har förmågan att direkt undersöka hur molekylära och miljömässiga faktorer påverkar ensemblemyosinkraftgenerering. Vidare har undersökning av kraftgenereringsmekanismer genom dessa aktin-myosinensembler potential att hjälpa till att modellera och förstå hur storskaliga cellulära uppgifter, såsom muskelkontraktion, sprider sig upp från molekylär nivå 9,10,13.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Etsning av täckglas
- Lös upp 100 g KOH i 300 ml 100% etanol i en 1,000 ml bägare. Rör om med en omrörningsstång tills majoriteten av KOH har upplösts.
VARNING: Koncentrerad KOH-lösning kan orsaka brännskador och skador på kläder. Använd handskar, ögonskydd och en labbrock. - Placera täckglas individuellt i rengöringsställ för täckglas.
OBS: Racks är utformade med slitsar som håller enstaka täckglas åtskilda för att möjliggöra etsning och sköljning på varje yta av täckglaset, dräneringshål i botten och gjorda av material som tål de hårda etsningsförhållandena. De kan skräddarsys eller köpas kommersiellt. - Förbered och märk tre 1 000 ml bägare: en med 300 ml etanol och två bägare med 300 ml vatten med omvänd osmos (RO).
OBS: Här hämtades RO-vatten från en laboratorievattenrenare, men det kan också köpas kommersiellt om en lokal renare inte är tillgänglig. - Placera var och en av de fyra bägarna i en badsonicator för att avgasa i 5 minuter.
- Sänk ner ett ställ med täckglas i bägaren av KOH och etanol och sonika i 5 min.
- Överför racken med täckglas från KOH/etanolbägaren till den enda etanolbägaren. Doppa rack upp och ner i bägaren tills det inte finns någon pärla.
OBS: Var noga med att inte störa täckglasen eller tappa racket kraftigt i bägaren. Detta kommer att orsaka att täckglasen kommer ut ur stället eller orsakar kemisk stänk. - Överför försiktigt stället med täckglas från etanolbägaren till en bägare med vatten, doppa upp och ner tills det inte finns någon pärla.
- Sänk ner stället med täckglas i bägaren av vatten som inte har använts ännu och sonika igen i 5 minuter.
- Använd en flaska för att spraya stället med täckglas med vatten tills det rinner av täckglasen smidigt. Upprepa med etanolen.
- Ställ racken för att torka i en ugn vid 90 °C i 20 min. Förvara racken med etsade täckglas vid rumstemperatur i slutna behållare för att förhindra kontaminering före användning.
2. Polymerisation av aktinfilament
- Gör lösning T
- I ett 50 ml koniskt rör tillsätt 3,94 g Tris-HCl och 0,147 g CaCl2. Tillsätt RO-vatten för att göra en total volym på 50 ml och blanda väl.
OBS: De slutliga koncentrationerna av lösning T är 500 mM Tris-HCl respektive 20 mMCaCl2 . - Märk röret Lösning T och förvara det vid 4 °C.
- I ett 50 ml koniskt rör tillsätt 3,94 g Tris-HCl och 0,147 g CaCl2. Tillsätt RO-vatten för att göra en total volym på 50 ml och blanda väl.
- Gör TC-buffert
- Blanda 40 ml RO-vatten och 1,5 ml lösning T i ett 50 ml koniskt rör. Ändra pH till 8,0 genom att tillsätta små mängder koncentrerad KOH. Tillsätt vatten för att göra 50 ml av lösningen och verifiera pH. Justera pH-värdet om det behövs.
OBS: Den slutliga TC-bufferten innehåller 5 mM Tris-HCl och 0,2 mMCaCl2 vid pH 8. - Märk röret TC och förvara det vid 4 °C.
- Blanda 40 ml RO-vatten och 1,5 ml lösning T i ett 50 ml koniskt rör. Ändra pH till 8,0 genom att tillsätta små mängder koncentrerad KOH. Tillsätt vatten för att göra 50 ml av lösningen och verifiera pH. Justera pH-värdet om det behövs.
- Gör FC-buffert
- Tillsätt 85 ml RO-vatten, 10 ml lösning T, 3,73 g KCl och 0,041 g MgCl2till en 100 ml buffertflaska. Ändra pH till 7,5 genom att lägga till små volymer koncentrerad KOH. Tillsätt vatten för att göra en slutlig volym på 100 ml och verifiera pH.
OBS: Den slutliga FC-bufferten innehåller 500 mM Tris-HCl, 500 mM KCl, 2 mM MgCl 2 och2 mM CaCl2vid pH 7,5. - Märk röret FC och förvara det vid 4 °C.
- Tillsätt 85 ml RO-vatten, 10 ml lösning T, 3,73 g KCl och 0,041 g MgCl2till en 100 ml buffertflaska. Ändra pH till 7,5 genom att lägga till små volymer koncentrerad KOH. Tillsätt vatten för att göra en slutlig volym på 100 ml och verifiera pH.
- Förbered allmän aktinbuffert (GAB).
- Blanda 485 μL TC-buffert, 10 μL 10 mM ATP och 5 μL 50 mM DTT i ett mikrocentrifugrör.
OBS: Slutliga buffertförhållanden är 5 mM Tris-HCl, 0,2 mM CaCl2, 0,5 mM DTT och0,2 mM ATP. - Märk den som GAB och förvara den vid 4 °C.
- Blanda 485 μL TC-buffert, 10 μL 10 mM ATP och 5 μL 50 mM DTT i ett mikrocentrifugrör.
- Förbered aktinpolymerisationsbuffert (APB).
- Blanda 455 μL FC-buffert, 25 μL 100 mM ATP och 20 μL 50 mM DTT i ett mikrocentrifugrör.
OBS: De slutliga buffertförhållandena är 50 mM Tris-HCl, 500 mM KCl, 2 mM MgCl 2, 2 mM CaCl 22 mM DTT och 5 mM ATP. - Märk tuben som APB och förvara den vid 4 °C.
- Blanda 455 μL FC-buffert, 25 μL 100 mM ATP och 20 μL 50 mM DTT i ett mikrocentrifugrör.
- Rekonstituera aktin
- Rekonstituera kaninskelettmuskelaktin genom att tillsätta 100 μL avjoniserat vatten till en 1 mg injektionsflaska med frystorkat aktin. Blanda väl genom att försiktigt pipettera upp och ner. Alikvotera i 5 μl prover, snäppfrys och förvara 10 mg/ml aktinalikvoter vid -80 °C.
- Rekonstituera biotinylerat kaninskelettmuskelaktin genom att tillsätta 20 μL RO-vatten. Alikvotera i 5 μl prover, snäppfrys och lagra 1 mg/ml biotinylerade aktinalikvoter vid -80 °C.
- Icke-märkt aktinpolymerisation med rhodaminfalloidinstabilisering
- Tina en injektionsflaska med 10 mg/ml aktin och förvara den på is.
- Förbered färsk GAB-buffert, tillsätt 100 μl GAB till aktinalikvoten och blanda genom att försiktigt pipettera upp och ner. Inkubera lösningen på is i 1 h.
- Förbered färsk APB under inkubationen. Efter inkubation polymerisera aktinet i filament genom att tillsätta 11 μL APB till aktinlösningen. Blanda väl genom att försiktigt pipettera upp och ner. Lägg på is i 20 min.
- Tillsätt 5 μL rhodaminmärkt falloidin till den nypolymeriserade aktinfilamentlösningen. Låt stå på is i mörkret i 1 timme.
- Förvara injektionsflaskan med rhodaminaktin insvept i aluminiumfolie i mörker vid 4 °C.
OBS: Det rekommenderas att använda dessa filament under högst 1 vecka. AF-kvalitet kan bekräftas varje dag genom en snabb avbildning av en flödescell som endast innehåller AF och tittar på konsekventa filament dag för dag.
- Biotinylerad aktinpolymerisation med Alexa Fluor 488 falloidinstabilisering
- Tina en injektionsflaska med 10 mg/ml aktin och 1 injektionsflaska med 1 mg/ml biotinylerat aktin och förvara dem på is.
- Gör färsk GAB-buffert.
- Kombinera de två injektionsflaskorna (steg 2.8.1) i ett 10:1-förhållande mellan aktin och biotinylerat aktin. Tillsätt 100 μl GAB till aktinblandningen och blanda väl genom att försiktigt pipettera upp och ner. Inkubera på is i 1 h.
- Gör färsk APB under inkubationen.
- Efter inkubationssteget polymerisera aktinet genom att tillsätta 11 μL APB till aktinlösningen. Blanda väl genom att pipettera upp och ner försiktigt. Inkubera på is i 20 min.
- Tillsätt 5 μL Alexa Fluor 488-märkt falloidin och inkubera på is i mörkret i 1 timme.
- Förvara den biotinylerade aktinflaskan insvept i aluminiumfolie i mörker vid 4 °C.
OBS: Dessa filament kan användas under högst 1 vecka.
3. Myosin och pärlberedning
- Rekonstituera Myosin II
- Snurra kort ner (~ 5 s) frystorkat skelettmyosin II för att samla det i botten av röret med hjälp av en standard minicentrifug.
- Rekonstituera myosinet till 10 mg/ml genom tillsats av 100 μl 1 mM DTT berett i RO-vatten.
- Späd stammyosinlösningen 10x genom att tillsätta 10 μl 10 mg/ml myosin till 90 μl 1 mM DTT i RO-vatten. Gör alikvoter med liten volym (1-5 μL), snäppfrys och förvara vid -80 °C.
OBS: Myosinaktivitet kan bekräftas genom att utföra en standardglidande filamentanalys som publicerats tidigare46,47. Se diskussionen för en kort beskrivning.
- Rengöring av streptavidinbelagda pärlor
- Späd 20 μl 1 μm streptavidinpärlor i 80 μl RO-vatten. Tvätta fyra gånger genom att spinna ner till 9 600 × g och rekonstituera i 100 μl RO-vatten.
- Sonicate i 2 min vid 40% amplitud och förvara de tvättade pärlorna på en rotator vid 4 °C.
4. Förberedelse av flödesceller
- Bered en poly-l-lysinlösning (PLL) genom att tillsätta 30 ml 100% etanol till ett 50 ml rör och tillsätt 200 μl 0,1% w / v poly-l-lysin i vatten och blanda väl.
- Tillsätt en etsad täckskiva i PLL-lösningen och låt den dra i 15 minuter. Ta bort täckglaset med pincett, var noga med att bara vidröra kanten på täckglaset när det dras upp från röret (se figur 1A-C). Ta tag i täckglasen vid kanterna med en handske.
- Torka täckglaset med ett filtrerat flygbolag tills det inte finns någon etanol kvar och inga rester på täckglaset.
- Applicera två bitar dubbelsidig tejp på mitten av ett mikroskopglas, 3-4 mm från varandra. Riv eller klipp av överflödig tejp som hänger av kanten på bilden.
- Lägg till den PLL-belagda täckglaset ovanpå tejpen vinkelrätt mot mikroskopglasets långa axel (bilda ett T) för att bilda en kanal.
- Använd ett litet rör för att komprimera täckglaset på tejpen och mikroskopet glider noggrant tills tejpen är transparent (bild 1A). Se till att det inte finns några bubblor i tejpen eftersom det kan orsaka läckage från flödeskanalen.
OBS: Flödescellen kan hålla en volym på 10-15 μL.
5. Beredning av aktomyosinbunt
- I separata rör späd varje typ av aktinfilament (rhodamin- och biotinylerad 488-märkt) 600x genom att blanda 0,5 μL av respektive, märkt aktin med 300 μL APB. Tillsätt ytterligare 5 μl av det motsvarande märkta falloidinet i varje rör och inkubera på is i mörkret i 15 minuter.
- Tillsätt ett syrerensningssystem med 1 μl beta-D-glukos vid 500 mg/ml till den biotinylerade aktinlösningen och tillsätt ett syrerensningssystem med 1 μl beta-D-glukos vid 500 mg/ml. Tillsätt 1 μl 100 mM ATP och 1 μl 100x utspädda, rengjorda streptavidinpärlor. Rör försiktigt med en pipettspets. Sätt suspensionen på en rotator vid 4 °C medan resten av aktomyosinbunten monteras.
- Tillsätt 15 μl av det utspädda rhodaminaktinet till PLL-flödescellen (figur 1D). Transportera överskottslösningen genom flödescellen men låt inte flödeskanalen bli torr. Inkubera i 10 minuter i en fuktighetskammare.
OBS: Fuktighetskamrar kan tillverkas av tomma pipettspetslådor med vatten tillsatt i botten och locket täckt av aluminiumfolie för att blockera ljus. - Bered en 1 mg/ml kaseinlösning i APB.
- Tillsätt 15 μl 1 mg/ml kasein för att förhindra ospecifik bindning av de efterföljande komponenterna (figur 1E). Inkubera i 5 minuter i en fuktighetskammare.
- Tillsätt önskad koncentration av myosin till det biotinylerade aktinet och pärlsuspensionen från steg 5.2. Rör försiktigt om med pipettspetsen och tillsätt sedan omedelbart 15 μL av steg 5.2-suspensionen + önskad myosinkoncentration till flödescellen (figur 1F,G). Inkubera i 20 min. Försegla de öppna ändarna av flödescellen med nagellack för att förhindra avdunstning under avbildning och optiska fångstexperiment.
OBS: En myosinlösningskoncentration på 1 μM ger robust buntning och kan användas som utgångspunkt för önskad anpassning av analysen (se figur 2).
6. Tvinga mätningar med optisk fälla (NT2 Nanotracker2)
OBS: Medan protokollet nedan är specifikt för NT2-systemet, kan denna analys användas med andra optiska fångstsystem, inklusive de som är specialbyggda, som också har fluorescensfunktioner. Det allmänna arbetsflödet förblir detsamma för att få bildens yta i fokus, utföra pärlkalibreringar och samla in data genom att hitta fluorescerande aktinbuntar. För NT2-systemet innehåller kompletterande figur S1, kompletterande figur S2, kompletterande figur S3, kompletterande figur S4, kompletterande figur S5, kompletterande figur S6 och kompletterande figur S7 uppgifter om det optiska svällningssystemet och programvarugränssnittet.
- Slå på kontrollboxen och lasern (kompletterande bild S1).
- Starta datorprogramvaran för optisk fälla genom att klicka på JPK Nanotracker-ikonen på skrivbordet.
- Väck fjärrkontrollen genom att klicka på Logitech-knappen i mitten (kompletterande bild S2).
- Slå på fluorescensmodulen genom att växla på / av-omkopplaren (kompletterande bild S3).
- Vrid filterkubtornet för ljusfältsavbildning (kompletterande bild S4).
- När systemet är klart, slå på lasern med laserströmknappen i skärmens vänstra nedre hörn till 50 mW och låt den stabiliseras i 30 minuter (kompletterande figur S5).
- Klicka sekventiellt på knapparna Belysning, Kamera, Mål och Scenrörelse i programvaran för att visa dessa fönster för visning och manipulation under experimentet. Slå på mikroskopbelysningen genom att klicka på På / Av-knappen och ställa in den på maximal effekt genom att klicka och dra fältet hela vägen till höger (kompletterande bild S5).
- Öppna provområdet och ta bort provhållaren från mikroskopsteget. Lägg till flödescellen, säkra den med metallprovhållarna och se till att bilden med täckglaset är på botten.
- Tillsätt 30 μl RO-vatten i mitten av bottenmålet. Låt inte pipettspetsen vidröra linsen. Sätt tillbaka provsteget igen.
OBS: Eftersom NT2-systemet använder ett vattensänkningsmål som fångstmål kan nedsänkningsmediet vara olika beroende på fångstmålet i användarens installation. - Höj det nedre målet med hjälp av kontrollpilarna på skärmen eller L2 på fjärrkontrollen tills vattenpärlan vidrör täckglaset (kompletterande figur S5).
- Sänk det övre målet tills ungefär hälften av avståndet till flödescellen har uppnåtts med pilarna på skärmen eller R2 på fjärrkontrollen. Tillsätt 170 μl RO-vatten till toppen av flödescellen direkt under det övre målet. Sänk det övre målet tills det bryter vattnets ytspänning och bildar en menisk.
- Flytta mikroskopsteget med pilplattan på fjärrkontrollen tills kanten på bandet intill flödeskanalen har uppnåtts. Stäng provdörren.
OBS: Ett "klick" när du stänger provdörren indikerar att laserluckan nu är öppen. Detta är en säkerhetsfunktion som endast tillåter slutaren att öppnas om dörren är stängd. - Använd fönstret Mål på skärmen och sätt bandets kant i fokus genom att föra upp det nedre målet med namnet Lasermål genom att klicka på den övre pilen med hjälp av kontrollerna på skärmen. Gör samma sak för det översta målet genom att klicka på den nedre pilen (kompletterande figur S5).
OBS: De dubbla pilarna flyttar målet eller scenen snabbare. Tejpens kant används för fokusering eftersom det är ett stort, lätt att hitta föremål som ligger nära täckplattans yta. Luftbubblor i tejpen är ett annat alternativ. Detta krävs dock inte om användaren har en automatiserad rutin för att hitta ytfokus eller en föredragen intern metod. - När tejpen är i fokus, stäng delvis irisen högst upp på den optiska fällan. Ta ner det övre målet tills irisens polygonform är synlig. Ta dessa kanter i fokus, öppna iris igen och koppla sedan ihop målen genom att klicka på hänglåsikonen (kompletterande figur S5).
- Hitta en flytande pärla och fånga den genom att klicka på Trap Shutter-knappen , som öppnar slutaren och låter fångstlasern träffa provet. Klicka på fällmarkören på skärmen och dra den för att flytta platsen för svällningslasern. När den är instängd, kalibrera pärlan för att korrelera spänningsmätningar till kraft och förskjutning.
- Klicka på kalibreringsknappen . Justera kalibreringsrutinen baserat på effektspektraanalys och anpassa hörnfrekvensen i programvaran för X-, Y- och Z-riktningarna (kompletterande figur S6).
- Klicka på Inställningar. Skriv in strängens diameter (1 000 nm) och skriv in temperaturen på scenen som finns längst ner till vänster i programvarufönstret. (se kompletterande figur S6).
- Klicka på Fälla 1. Klicka på X Signal. Klicka på Kör för att utföra hörnfrekvenspassningen. Klicka och dra i fönstret för att optimera funktionspassningen. Klicka på Använd den för känslighets- och styvhetsvärden. Klicka på Acceptera värden. Upprepa för Y- och Z-signalerna. Stäng fönstret. (se kompletterande figur S6).
OBS: Pärlkalibreringsrutiner på andra optiska svällningssystem eller specialbyggda system som har testats robust av användaren, såsom ekvipartitionsmetoden eller dragkraftsmetoden, är också acceptabla57,58. - Hitta en aktomyosinbunt genom att söka efter pärlor bundna till AFs på ytan av täckglaset.
- När en pärla som inte är full av andra flytande pärlor detekteras, observera AF: erna runt den genom fluorescensavbildning för att verifiera närvaron av en bunt.
- Kontrollera att det finns ett paket genom att leta efter båda fluorescerande AF: erna samlokaliserade. Slå på den vita ljuskällan och använd lämplig filterkub för att avbilda varje aktinfilament genom att vrida tornet (488 nm och 532 nm excitationsfilterkuber för Alexa Fluor 488 respektive rhodaminexcitation). Se kompletterande figur S4.
OBS: Ett kontrollexperiment för att verifiera fluorescensintensiteten hos enskilda AF kan vara användbart för att identifiera buntar som består av en enda 488- och enda rhodaminmärkta filament, eller tillämpliga på vilken uppsättning fluoroforer användaren väljer att använda. - När du har verifierat fångar du pärlan som är fäst vid buntens övre filament genom att klicka på Trap Shutter-knappen .
- Använd kontrollerna på skärmen för att spela in data genom att klicka på oscilloskopknappen (kompletterande figur S7). För att visualisera mätningar utan att spela in data, klicka på Start. För att spara all data, klicka på Autosave. För att spela in mätningar, klicka på Starta inspelning. Välj vilka data som ska visualiseras i realtid (position, kraft, x-riktning, y-riktning) genom att välja från rullgardinsmenyn X-signal eller Y-signal. Kom ihåg att xdirection är vänster till höger och y-riktning är upp och ner på skärmen. Se kompletterande figur S7.
OBS: Data sparas som .out-filer och inkluderar tid, spänning, förskjutning och kraft för varje riktning. Dessa filer kan exporteras till annan programvara för visualisering och analys.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Flödesceller som innehåller aktomyosinbuntsystemen är av standardutförande, bestående av ett mikroskopglas och ett etsat täckglas åtskilt av en kanal gjord av dubbelsidig tejp (figur 1). Analysen byggs sedan från täckglaset upp med hjälp av iscensatta introduktioner som beskrivs i protokollet. Den slutliga analysen består av mall rhodaminmärkta aktinfilament; Den önskade myosinkoncentrationen (1 μM användes för de representativa resultaten i figur 2 och figur 3). biotinylerade, Alexa Fluor 488-märkta aktinfilament; 1 μm streptavidinpärlor; syrerensningssystemet; ATP; och APB-buffert. Flera buntar kommer att bildas per flödescell, och de aktinkoncentrationer som beskrivs ovan ger tillräckligt avstånd mellan buntar för att säkerställa inga oönskade interaktioner. Detta underlättar också att få flera kraftmätningar per flödescell för att öka datainsamlingseffektiviteten. Kraftprofiler ska kunna reproduceras i en flödescell och från flödescell till flödescell.
Medan protokollet ovan är inriktat på användningen av en kommersiell optisk fångstinställning, kan flödescellen och analysen som presenteras här enkelt användas för ett annat kommersiellt instrument eller specialbyggd optisk fångstinställning i kombination med ett mikroskop eller mikroskopsteg och har fluorescensavbildningsfunktioner. När alla flödescellsadditioner är klara enligt ovanstående protokoll är aktomyosinbuntarna på bilden (figur 1) redo för omedelbar mätning. Flödescellen läggs till det optiska fällmikroskopsteget, flera pärlkalibreringsmätningar förvärvas och buntar identifieras genom fluorescenskolokalisering av buntfilamenten. En pärla bunden till en bunt fångas och förskjutningen och motsvarande kraftmätning börjar. Användaren kan observera förvärvet av data i realtid på datorskärmen. Beroende på koncentrationen av myosin som används i flödescellen kan bunten börja uppvisa betydande rörelse omedelbart, eller det kan ta 30 s-1 min att effektivt se en ökning av förskjutning / kraft.
Ett representativt kraftspår visas i figur 3A där myosinmotorerna uppvisar en stadig ramp i kraft följt av en platå. Det är typiskt att se dessa typer av spår utvecklas under 2-5 min. Det är emellertid också möjligt att mäta aktomyosinbuntar som inte genererar någon nettokraft (figur 3B). Dessa spår uppträder som baslinjebrus eller uppvisar ingen betydande nettoökning i kraft över 90 s. Detta beror sannolikt på en låg lokal koncentration av motor som inte tillåter produktiv glidning, eller bunten är i en ogynnsam parallell orientering där filamentens plus- och minusändar är inriktade.
Eftersom innehållet i flödescellen kan vara mottagligt för nedbrytning från den infallande belysnings- och fångstlasern, lokal uppvärmning på bilden över tid och generering av radikala syrearter, rekommenderas det starkt att inte använda samma flödescell i mer än 1 h. För maximal effektivitet föreslås att en annan analys inkuberas när data samlas in. Förskjutnings-/kraftspårning kan exporteras från den optiska svällningsprogramvaran till Excel, Matlab, Igor eller andra datahanteringsprogram för ytterligare filtrering och analys. Data som kan extraheras från sådana optiska fångstensemble/buntexperiment inkluderar olika typer av kraftgenereringsprofiler (baslinje, ramp/platå) under varierande analysförhållanden, hastighet för kraftgenerering, maximal kraftgenerering, ensemblekinetiskt och stegbeteende genom stegstorlekar och uppehållstider mellan steg eller steg av steg, samt pliktförhållande. Användaren kan också ändra analysförhållandena för att jämföra hur tillsats av olika typer av myosinmotorer, tillsats av aktinbindande proteiner eller förändrade buffertförhållanden påverkar dessa ensemblekraftgenereringsegenskaper.
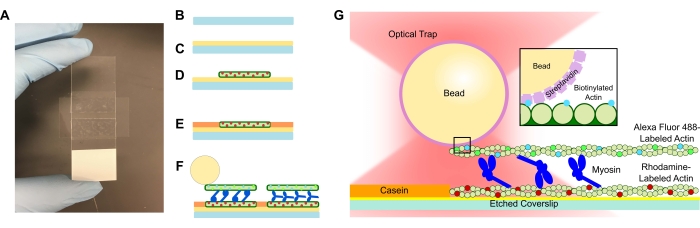
Figur 1: Analysschema. (A-C) Etsade täckglas är belagda med poly-L-lysin och används för att bilda flödescellen med hjälp av dubbelsidig tejp och ett mikroskopglas. Tidsinställda introduktioner och inkubationssteg som beskrivs i protokollet resulterar i rhodaminmärkt falloidinstabiliserat aktin som mall eller bottenfilament (D), följt av kaseinblockering för att förhindra ospecifik bindning (E), och (F) Alexa Fluor 488 falloidinstabiliserat biotinylerat aktin som last- eller toppfilament, och team av myosin II som skjuter isär filamenten och genererar kraft när ATP införs. Motorernas geometri och tvärbindningens natur i bunten kan variera under olika förhållanden, såsom saltkoncentration59. Tidigare studier har visat att myosinsvansdomänen har förmågan att interagera med aktinfilament och långsam ensemblemotilitet46. Myosinhuvuden i tunga meromyosinexperiment visar emellertid bindning av varje huvud till intilliggande aktinfilament60. (G) Streptavidinpärlor används som det optiska handtaget för fällan och binder endast till lastens biotinylerade aktinfilament, vilket hjälper till att validera att lämpliga buntar bildas på bilden. Klicka här för att se en större version av denna siffra.

Figur 2: Fluorescerande aktomyosinbuntar. Fyra olika möten mellan aktinfilament och buntar i buntanalysen som presenteras i figur 1. Den övre lasten biotinylerad aktinfilament med Alexa Fluor 488 phalloidinkanal visas till vänster och den nedre mallen aktinfilament med rhodaminfalloidinkanalen är till höger. Längst ner visas samma figur med färgade linjer överlagrade för att hjälpa till att styra ögat. (A) Ett övre aktinfilament finns nära ett bottenaktinfilament men har ett ofullständigt överlägg. Detta skulle inte användas för buntexperiment. (B) Övre och nedre aktinfilament är samlokaliserade, och intensiteten hos varje filament bekräftar att de är var och en av filamenten i bunten. Detta skulle vara en bra kandidat för buntexperiment. (C) En stor bunt självmonterade rhodaminfilament finns på botten. Även om det finns en motsvarande toppaktinfilament som är samlokaliserad, finns det för många bottenfilament närvarande; Således skulle den inte användas för buntexperiment. Detta är också ett exempel på hur fluorescensintensiteten ökar när flera aktinfilament av samma typ buntas. Användaren kan använda detta som en mätare för att bedöma enstaka filament kontra buntar av samma filamenttyp. (D) Det finns ett bottenfilament utan motsvarande toppfilament, vilket också bekräftar att det inte finns någon genomblödning. Detta skulle inte användas för buntexperiment. Vi noterar att glödtrådarnas intensitet i Alexa Fluor 488-kanalen är låg och tror att det beror på filteruppsättningen som används (Filteruppsättning 09 från Zeiss). Filteruppsättningen som används för rhodaminkanalen är Filter Set 43 från Zeiss. Klicka här för att se en större version av denna siffra.

Figur 3: Myosin II ensemblekraftgenerering. Representativa spår av skelettmyosin II-motorer som genererar kraft inom den konstruerade in vitro-aktinstrukturhierarkin. Myosinmotorerna arbetar tillsammans för att kollektivt och produktivt generera kraft tills en platå uppnås och kraften upprätthålls (A) eller upplever antagonisering nära baslinjen (B). Klicka här för att se en större version av denna siffra.
Kompletterande figur S1: Bruker/JPK Nanotracker2 optisk fälla. (A) Datorskärm. (B) Datorns tangentbord. (C) Datortorn. (D) Kontrollbox. (E) Laser strömförsörjning. (F) Optisk fälloptiklåda. (G) Inverterat mikroskop. (H) Dörr till mikroskopstadium. (I) Polarisatorreglage för att växla mellan ljusfält och differentiell interferenskontrastavbildning. Klicka här för att ladda ner den här filen.
Kompletterande bild S2: Fjärrkontroll för optisk fälla. (A) Knappsats för att placera det motoriserade steget. (B–C) Justera svällningspositionen. (D) A, X och B slår på och av huvudluckan, fälla 1 slutare respektive fälla 2 slutare. (E) Logitech-knappen används för att väcka handkontrollen. (F) Upp- och nedknapparna som används för att placera svällningsmålet. (G) Upp- och nedknapparna som används för att placera detekteringsmålet. Observera att fjärrkontrollen inte krävs, och alla dessa manipuleringar kan utföras i programvaran. Det är dock bekvämt att kunna styra målen och scenpositionen medan man tittar på mikroskopscenen. Klicka här för att ladda ner den här filen.
Kompletterande figur S3: Fluorescensmodul för optisk fälla. Den vita ljuskällan 89North PhotoFluorescens är kopplad till baksidan av det inverterade mikroskopet. Den slås på och av med en vippbrytare (pil). Klicka här för att ladda ner den här filen.
Kompletterande figur S4: Fluorescensfilterkubtorn. Tornet (pilen) kan vridas för att använda filterkuben som är nödvändig för avbildning i DIC-, rhodamin- eller Alexa Fluor 488-färgämnen. Observera att filterkuber kan bytas ut för att anpassa inställningen för att använda olika fluoroforer. Klicka här för att ladda ner den här filen.
Kompletterande figur S5: Nanotracker2-programvara. (A) Laserströmbrytare och kontroll. (B) Objektiv placeringsfönster. Riktningspilar används för att flytta identifieringsmålen (överst) och svällningsmålen (nederst). Dubbla pilar flyttar målen med högre hastighet. Den blå och röda knappen längst ned till vänster kopplar bort målen och drar tillbaka dem till sin ursprungliga position. Detta är nödvändigt för när du tar prover in och ut ur mikroskopstadiet. Den tredje knappen från vänster med målen och hänglåsikonen "kopplar" målen så att när de både är i fokus och uppnår Kohler-belysning kan användaren flytta både fångst- och detekteringsmålen upp och ner i z-axeln. (C) Provpositioneringsfönster som används för att flytta mikroskopsteget i x- och y-axeln. Dubbla pilar flyttar scenen med högre hastighet. Det här fönstret aktiveras genom att klicka på pilikonen upp / ner och vänster / höger på toppmenyn. (D) Fönstret för kameravisualisering. Skiftnyckelikonen kan användas för att ställa in anpassade bildförhållanden. Det här fönstret aktiveras genom att klicka på kameraikonen i toppmenyn. (E) Fönster för mikroskopbelysning. Det här fönstret aktiveras genom att klicka på lampikonen i toppmenyn. Klicka här för att ladda ner den här filen.
Kompletterande bild S6: Kalibreringsfönster. a) Detta fönster används för pärlkalibrering och aktiveras genom att klicka på Cal-ikonen på toppmenyn. För att kalibrera en pärla uppnås en bästa passform av hörnfrekvensen i x-, y- och z-signalerna. (B) För varje signal, välj lämplig signalknapp längst upp till vänster. (C) Klicka på kör och optimera passformen genom att klicka och dra i det gröna fönstret (D). (E) När du är nöjd med passformen klickar du på Använd den för känslighet och styvhet. Detta möjliggör inspelning av förskjutning i nanometer och kraft i piconewtons. (F) Klicka sedan på Acceptera värden längst ner till vänster. Upprepa för y- och z-riktningarna. Klicka här för att ladda ner den här filen.
Kompletterande figur S7: Fönstret Datainsamling. Detta fönster används för att hämta positions- och kraftdata och låter användaren se mätningarna i realtid. (A) Det här fönstret aktiveras genom att klicka på x, t-ikonen på toppmenyn. (B) Användaren kan växla mellan att visa x- och y-signalerna. (C) Klicka på Start för att börja visualisera data. Klicka på Autosave för att spara data. Klicka på Starta inspelning för att börja spela in och spara data. Klicka här för att ladda ner den här filen.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
En in vitro-studie med optisk pincett i kombination med fluorescensavbildning utfördes för att undersöka dynamiken hos myosinensembler som interagerar med aktinfilament. Actin-myosin-aktinbuntar monterades med användning av muskelmyosin II, rhodaminaktin i botten av bunten och på täckytan och 488-märkta, biotinylerade aktinfilament på toppen av bunten. Aktinprotein från kaninmuskeln polymeriserades och stabiliserades med användning av allmänna aktinbuffertar (GAB) och aktinpolymeriserande buffertar (APB). GAB och APB måste vara nyberedda varje dag i labbet med ATP, FC-buffert och TC-buffert. Muskelmyosin II användes för att bilda aktin-myosin-aktinsmörgåsarna. Falloidin användes för fluorescerande färgning av aktinfilamenten, såväl som stabilisering in vitro.
Myosinaktivitet kan bekräftas genom att utföra en standardglidande filamentanalys som tidigare publicerats46,47. Myosin II och dess subfragment kan binda till täckytan i olika riktningar, och närvaron av svansdomänen kan sakta ner filamentglidningen jämfört med analyser med tungt meromyosin46,48,49. Glidning och ytrörelse kan dock fortfarande observeras. En tydligare demonstration av myosinaktivitet är aktiv aktinfilamentbrytning som kan observeras där längre aktinfilament bryts upp i mindre fragment som sedan glider bort i flera riktningar. Detta sker på grund av den höga koncentrationen av aktiva motorer på ytan, har observerats av flera laboratorier och sker inte utan aktiva myosinmotorer närvarande 42,50,51,52,53,54. Vidare hjälper buntanalysen som presenteras här att lindra rörlighetsproblem som främst har associerats med glidfilamentanalysen, såsom de olika motorbindningsorienteringarna på en glasbeläggning, eftersom buntanalysen involverar kaseinblockering av glasytan så att motorer binder inom bunten 47,55,56.
Det första steget är att lägga till rhodaminaktinfilament som botten eller mallfilament till en poly-L-lysinbelagd täckslip i en flödescell. Poly-L-lysin används för att främja aktinbindning eftersom poly-lysin är positivt laddat medan aktin har negativa laddningar och har använts i tidigare cytoskeletala in vitro-analyspreparat61,62,63. Före buntbildning tillsattes olika aktinutspädningar till en flödescell för att optimera aktinkoncentrationen. I detta fall var 600x från beståndet den optimala utspädningen som gav ett tillräckligt antal mallfilament för buntbildning men med tillräckligt avstånd så att buntarna individualiserades. Utspädning utfördes med användning av APB-bufferten. Tillsats av rhodaminaktin följdes av ett lager kasein för att blockera ytan och undvika ospecifik bindning. Flödescellen inkuberades i 30 minuter och tvättades efter inkubation med buffert för att tvätta bort eventuella obundna aktinfilament. Slutligen tillsattes en kombination av myosin, 488/biotinaktin och streptavidinbelagda pärlor till flödescellen för att underlätta bildandet av aktin-myosinbuntar. Pärlkoncentrationen bör vara sådan att det finns tillräckligt för att binda ytbundna buntar och tillräckligt i suspension för att underlätta kalibrering. För hög pärlkoncentration kan dock orsaka svårigheter under fångstexperiment på grund av att närliggande pärlor faller i laserfällan och stör mätningen. Myosinmotorer tillsätts till kombinationen precis innan de injiceras i bilden så att myosinmotorerna inte i förebyggande syfte aggregeras med lasten eller det övre biotinylerade aktinfilamentet och därmed binder botten rhodamin för att bunta biotinylerade aktinfilament.
NT2 optiskt fångstsystem är en kommersiell optisk fälla med kombinerade ljusfält, differentiell interferenskontrast (DIC) och epifluorescensavbildningsmetoder. Den är kopplad till ett Zeiss AxioObserver 3 inverterat mikroskop med 100x / NA 1.46 och 63x / NA 1.0 vattensänkningsfångning och detektionsmål. Systemet är utrustat med klick- och dra-fångstfunktioner för en laserfälla och kan användas vid avbildning i någon av de listade metoderna tidigare. De bildade buntarna detekteras och bekräftas med hjälp av fluorescensavbildning. Att ha en vit ljuskälla med lämpliga filterkuber (GFP/FITC och TRITC/CY3) möjliggör snabb växling mellan filamentavbildning. Kolokaliserade AF verifierades genom att visualisera AF: erna vid de olika excitationsvåglängderna innan de tog varje kraftmätning med optisk pincett. Eftersom filamenten kan fotoblekas snabbt även med ett syrerensningsreagens, föreslås det att forskare optimerar visualiseringsparametrar som intensitet och exponeringstid innan de utför buntexperimenten.
Optisk fångst användes för att göra kraftmätningarna med hjälp av streptavidinpärlorna i närvaro av ATP för att binda det biotinylerade lastaktinfilamentet och aktivera myosinkraftgenerering som en kraftgivare. Förskjutnings- och kraft- kontra tidsdata som erhållits genom optisk svällning extraherades från svällningsprogramvaran för analys. Men den kommersiella fångstprogramvaran tillhandahåller också analysrutiner som kan användas, eller anpassade algoritmer i andra program kan programmeras av användaren för att visualisera och analysera svällningsdata. På anpassade optiska svällningssystem kan användaren ha excitationslasrar istället för en vit ljuskälla med filter, som också är acceptabla att använda. Vidare kan fluorescerande färgämnen ändras för att passa den befintliga utrustning som en användare kan ha om emissionsspektra inte överlappar varandra och orsakar blödning.
Vi noterar att analysen som presenteras är en baslinjeanalys som kan anpassas ytterligare av användaren beroende på deras forskningsfråga inom aktomyosinensemblemekaniken. Det allmänna arbetsflödet kan också tillämpas på andra in vitro-cytoskelettensemblesystem som kan vara av intresse, såsom mikrotubulibuntanalyser som bildar minimala modeller av mitotisk spindel 32,61,63,64,65,66. Ändringar kan inkludera men är inte begränsade till att ändra fluoroforetiketterna som passar användarens befintliga installation; ändra myosinkoncentration, konstruktion eller isotyp; och titrering av buffertförhållanden, bland andra aspekter.
Potentiella utmaningar är möjliga när du utför denna analys. Vid bildandet av aktin-myosinbuntarna kan myosinkoncentrationen i aktinbuntarna inte vara homogen över bilden. För att tillgodose detta kommer flera buntar över hela bilden att mätas för att säkerställa att motorfördelnings- och kraftgenereringsprofiler samplas korrekt. Det är också utmanande att känna till buntorientering om detta krävs för tolkning av kraftdata. Således bör flera försök tas för varje bunt. Man kan också införliva aktinfilamentändmärkning genom fluorescerande gelsolin eller gelsolinbelagda pärlor av mindre storlek än det optiska fångsthandtaget. Fluorescensavbildning kan också användas för att titta på x- och y-komponentkrafter för att härleda buntorientering. Eftersom myosinaggregeringstillståndet i hög grad påverkas av buffertens jonstyrka med bildning av tjocka filament som sker vid snabb utspädning av KCl, bör buffertsaltkoncentrationen övervakas på lämpligt sätt67,68.
Tidigare studier som använde andra in vitro-metoder som glidanalyser var till hjälp för att identifiera myosindomänernas roll och studera konfigurationen och interaktionerna mellan myosin och andra aktinbindande proteiner. Dessa metoder har emellertid en nackdel genom att bindning av myosin på en styv yta kommer att begränsa potentialen för samordning mellan myosinmotorer och därmed mekanosenserande återkoppling som uppstår för att avgöra om motorensemblen är i ett högt eller lågt arbetsförhållandeläge 33,35,41,69. Vidare ger optisk fångst med enkelmyosinmotornätverk inte en klar förståelse för hur myosinmotorer interagerar med varandra och med aktinfilament. Protokollet som utvecklats här möjliggör undersökning av myosinmotorensembledynamik inom ett kompatibelt, hierarkiskt aktinnätverk. Det är också anpassningsbart när det gäller motorfilamentensembleegenskaper som koncentration, isoform och buffertmiljö, bland andra aspekter, för att möjliggöra systematisk undersökning. Det presenterade protokollet är en plattform för framtida studier av mer komplexa aktomyosinnätverk och upprätthåller precisionen i förskjutnings- och kraftgenereringsmätningar som underlättas av optisk fångst som traditionellt har använts för enmolekylstudier.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
Författarna har inga intressekonflikter att deklarera.
Acknowledgments
Detta arbete stöds delvis av University of Mississippi Graduate Student Council Research Fellowship (OA), University of Mississippi Sally McDonnell-Barksdale Honors College (JCW, JER), Mississippi Space Grant Consortium under bidragsnummer NNX15AH78H (JCW, DNR) och American Heart Association under bidragsnummer 848586 (DNR).
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
