

Summary
विट्रो में एक्टोमायोसिन बंडलों का गठन और ऑप्टिकल ट्वीज़र्स का उपयोग करके मायोसिन एन्सेंबल फोर्स जनरेशन को मापना प्रस्तुत और चर्चा की गई है।
Abstract
मायोसिन मोटर प्रोटीन हैं जो एटीपी को एक्टिन फिलामेंट (एएफ) पटरियों के साथ कदम उठाने के लिए हाइड्रोलाइज करते हैं और गतिशीलता और मांसपेशियों के संकुचन जैसी सेलुलर प्रक्रियाओं में आवश्यक हैं। उनके बल-उत्पादन तंत्र को समझने के लिए, मायोसिन II की जांच एकल-अणु (एसएम) स्तर पर और ऑप्टिकल ट्रैपिंग जैसे बायोफिज़िकल तरीकों का उपयोग करके विट्रो में मोटर्स की टीमों के रूप में की गई है।
इन अध्ययनों से पता चला है कि मायोसिन बल पैदा करने वाला व्यवहार तीन-मोती व्यवस्था में एकल-अणु स्तर से एक कठोर मोती या कवरस्लिप सतह पर एक साथ काम करने वाले मोटर्स के समूहों में स्थानांतरित होने पर बहुत भिन्न हो सकता है। हालांकि, ये परख निर्माण विस्कोस्टिक संरचनात्मक पदानुक्रम के भीतर मायोसिन की समूह गतिशीलता का मूल्यांकन करने की अनुमति नहीं देते हैं क्योंकि वे एक सेल के भीतर होंगे। हमने कई एक्टिन फिलामेंट्स के साथ बातचीत करने वाले मायोसिन एन्सेंबल द्वारा बल उत्पादन के यांत्रिकी की जांच करने के लिए ऑप्टिकल ट्वीज़र्स का उपयोग करके एक विधि विकसित की है।
ये एक्टोमायोसिन बंडल एक पदानुक्रमित और अनुरूप वातावरण में जांच की सुविधा प्रदान करते हैं जो मोटर संचार और एन्सेंबल फोर्स आउटपुट को पकड़ता है। परख की अनुकूलन योग्य प्रकृति प्रयोगात्मक स्थितियों को बदलने की अनुमति देती है ताकि यह समझा जा सके कि मायोसिन पहनावा, एक्टिन फिलामेंट बंडल, या आसपास के वातावरण में संशोधन के परिणामस्वरूप अलग-अलग बल आउटपुट होते हैं।
Introduction
मोटर प्रोटीन जीवन के लिए आवश्यक हैं, रासायनिक ऊर्जा को यांत्रिक कार्य में परिवर्तित करते हैं 1,2,3. मायोसिन मोटर्स एक ट्रैक के समान फिलामेंट्स के साथ कदम उठाकर एक्टिन फिलामेंट्स के साथ बातचीत करते हैं, और एक्टिन-मायोसिन नेटवर्क की गतिशीलता मांसपेशियों के संकुचन, सेल गतिशीलता, साइटोकिनेसिस के दौरान सिकुड़ा हुआ रिंग और सेल के अंदर कार्गो की आवाजाही को अन्य आवश्यक कार्यों के बीच अंजाम देती है 3,4,5,6,7,8 . चूंकि मायोसिन की कई आवश्यक भूमिकाएं हैं, इसलिए मायोसिन-एक्टिन नेटवर्क की कार्यक्षमता में विफलता से रोग का विकास हो सकता है, जैसे कि मायोसिन भारी श्रृंखला में उत्परिवर्तन जो हाइपरट्रॉफिक कार्डियोमायोपैथी (एचसीएम) 9,10,11,12,13,14 में हृदय हाइपरकॉन्ट्रेसिटी का कारण बनता है . मांसपेशियों के संकुचन में, व्यक्तिगत मायोसिन मोटर्स आवश्यक यांत्रिक ऊर्जा प्रदान करने के लिए एक पहनावा के रूप में काम करके एक दूसरे के साथ सहयोग करते हैं जो एएफ 4,15,16,17,18 के सापेक्ष स्लाइडिंग को अंजाम देता है। मायोसिन मोटर्स एएफ के बीच क्रॉसब्रिज बनाते हैं और अपने मेकेनोकेमिकल चक्र के कारण विरूपण परिवर्तनों का उपयोग करते हैं ताकि सामूहिक रूप से संरेखित फिलामेंट्स17,18,19,20,21 के कांटेदार छोर की ओर बढ़ सकें।
ऑप्टिकल ट्रैपिंग जैसी तकनीकों का उपयोग करके एसएम स्तर पर मात्रात्मक इन विट्रो गतिशीलता परख के विकास ने एसएम बल उत्पादन और चरण आकार 22,23,24,25,26,27,28,29,30 को मापने सहित व्यक्तिगत मायोसिन मोटर्स के कार्य के बारे में अभूतपूर्व विवरण एकत्र करने की सुविधा प्रदान की है। . फाइनर एट अल ने एकल मायोसिन II मोटर्स23,31 के बल-पीढ़ी यांत्रिकी की जांच के लिए "तीन-मोती" या "डंबल" ऑप्टिकल ट्रैपिंग परख विकसित की। चूंकि मांसपेशी मायोसिन II एएफ अनुबंध करने के लिए टीमों में काम करता है, लेकिन एसएम स्तर पर गैर-प्रक्रियात्मक है, ऑप्टिकल ट्रैपिंग परख अभिविन्यास को क्लासिक मोटर-बाउंड बीड दृष्टिकोण32 से पुनर्व्यवस्थित किया जाना था। डंबल परख बनाने के लिए, दो ऑप्टिकल ट्रैप का उपयोग एक कवरस्लिप-संलग्न मोती से बंधे मायोसिन मोटर पर एएफ रखने के लिए किया गया था, और एकल मोटर द्वारा बल उत्पादन को ट्रैप23 के भीतर एएफ के आंदोलनों के माध्यम से मापा गया था।
हालांकि, एसएम बल और एकल मोटर / एकल फिलामेंट परख अभिविन्यास का उपयोग सिस्टम-स्तरीय बल उत्पादन के बारे में पूरी छवि नहीं देता है क्योंकि मायोसिन II सहित कई मोटर प्रोटीन अलगाव में काम नहीं करते हैं और अक्सर अपने भागों के योग के रूप में कार्य नहीं करते हैं 15,16,17,32,33,34,35,36 . अधिक जटिल संरचनाएं जिनमें एक से अधिक फिलामेंट के साथ बातचीत करने वाले एक से अधिक मोटर शामिल हैं, मायोसिन और एक्टिन फिलामेंट्स के नेटवर्क15,32 के तालमेल को बेहतर ढंग से समझने के लिए आवश्यक हैं। डंबल परख अभिविन्यास का उपयोग एक मोती से जुड़े कई मायोसिन या सतह से जुड़े मायोसिन-मोटी फिलामेंट का उपयोग करके छोटे पहनावा बल उत्पादन की जांच करने के लिए किया गया है और मोटर्स को निलंबित एएफ 4,23,34,37,38,39,40 के साथ बातचीत करने की अनुमति देता है।
अन्य छोटे समूहों में एक इन विट्रो फिलामेंट ग्लाइडिंग परख शामिल है जिसमें मायोसिन मोटर्स को कवरस्लिप सतह पर लेपित किया जाता है, और एएफ से बंधे एक मोती का उपयोग मोटर्स 4,35,36,38,39,40,41,42,43 की टीम द्वारा उत्पन्न बल की जांच के लिए किया जाता है। . इन दोनों मामलों में, मायोसिन एक कठोर सतह से बंधे होते हैं - मोती या कवरस्लिप - और एक एएफ का उपयोग करते हैं। इन मामलों में, मोटर्स स्वतंत्र रूप से स्थानांतरित करने या एक-दूसरे के साथ संवाद करने में सक्षम नहीं होते हैं, न ही मायोसिन को कठोर रूप से बाध्य करने से अनुपालन, पदानुक्रमित वातावरण परिलक्षित होता है जिसमें मोटर्स सरकोमेरे32 में एक साथ काम करेंगे। पिछले अध्ययनों ने सुझाव दिया है कि मायोसिन II अपने पर्यावरण को समझ सकता है और बल उत्पादन और कर्तव्य अनुपात41,44,45 जैसी विशेषताओं को बदलकर विस्कोस्टिक या मोटर एकाग्रता स्थितियों को बदलने के लिए तदनुसार अनुकूलित कर सकता है। इस प्रकार, एक ऑप्टिकल ट्रैपिंग परख विकसित करने की आवश्यकता है जो मायोसिन द्वितीय एन्सेंबल फोर्स जनरेशन के यंत्रवत आधार की अधिक यथार्थवादी तस्वीर को चित्रित करने के लिए मोटर संचार और सिस्टम कॉम्प्लीेंसी को बढ़ावा देता है और कैप्चर करता है।
यहां, हमने एक्टोमायोसिन बंडल या सैंडविच बनाकर ऑप्टिकल ट्रैपिंग के साथ विट्रो में पदानुक्रमित संरचना को जोड़ने के लिए एक विधि विकसित की, जिसमें दो एक्टिन फिलामेंट्स के बीच बातचीत करने वाले कई मायोसिन मोटर शामिल थे। इस मॉड्यूलर परख ज्यामिति में सीधे जांच करने की क्षमता है कि आणविक और पर्यावरणीय कारक कैसे मायोसिन बल उत्पादन को प्रभावित करते हैं। इसके अलावा, इन एक्टिन-मायोसिन एन्सेंबल के माध्यम से बल उत्पादन तंत्र की जांच करने से मॉडलिंग और समझने में सहायता करने की क्षमता है कि मांसपेशियों के संकुचन जैसे बड़े पैमाने पर सेलुलर कार्य आणविक स्तर 9,10,13 से कैसे फैलते हैं।
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. नक़्क़ाशी कवरलिप्स
- 1,000 एमएल बीकर में 100% इथेनॉल के 300 एमएल में 100 ग्राम केओएच को भंग करें। एक हलचल पट्टी के साथ हिलाएं जब तक कि कोह का अधिकांश हिस्सा भंग न हो जाए।
सावधानी: केंद्रित कोह समाधान कपड़ों को जलने और नुकसान पहुंचा सकता है। दस्ताने, आंखों की सुरक्षा और एक प्रयोगशाला कोट पहनें। - कवरस्लिप सफाई रैक में व्यक्तिगत रूप से कवरलिप रखें।
नोट: रैक को स्लिट्स के साथ डिज़ाइन किया गया है जो सिंगल कवरलिप्स को अलग-अलग रखते हैं ताकि कवरस्लिप के प्रत्येक चेहरे पर नक़्क़ाशी और कुल्ला किया जा सके, तल में छेद निकाला जा सके, और ऐसी सामग्री से बना हो जो कठोर नक़्क़ाशी की स्थिति का सामना कर सके। उन्हें कस्टम-निर्मित या व्यावसायिक रूप से खरीदा जा सकता है। - तीन 1,000 एमएल बीकर तैयार करें और लेबल करें: एक 300 एमएल इथेनॉल के साथ और दो बीकर 300 एमएल रिवर्स ऑस्मोसिस (आरओ) पानी के साथ।
नोट: यहां, आरओ का पानी एक प्रयोगशाला जल शोधक से प्राप्त किया गया था, लेकिन यदि स्थानीय प्यूरीफायर उपलब्ध नहीं है तो इसे व्यावसायिक रूप से भी खरीदा जा सकता है। - चार बीकरों में से प्रत्येक को स्नान सोनिकेटर में 5 मिनट के लिए रखें।
- कोह और इथेनॉल के बीकर में कवरलिप्स का एक रैक डुबोएं और 5 मिनट के लिए सोनिकेट करें।
- इथेनॉल बीकर से कवरलिप्स के रैक को इथेनॉल-ओनली बीकर में स्थानांतरित करें। बीकर में ऊपर और नीचे डुबोएं जब तक कि कोई मोती न हो।
नोट: ध्यान रखें कि कवरलिप्स को परेशान न करें या रैक को बीकर में जबरदस्ती न गिराएं। इससे कवरलिप्स रैक से बाहर आ जाएंगे या रासायनिक छींटे पड़ेंगे। - इथेनॉल बीकर से कवरलिप्स के रैक को सावधानीपूर्वक पानी के बीकर में स्थानांतरित करें, जब तक कि कोई मोती न हो, ऊपर और नीचे डुबकी लगाएं।
- कवरलिप्स के रैक को पानी के बीकर में डुबोएं जिसका अभी तक उपयोग नहीं किया गया है और 5 मिनट के लिए फिर से सोनिकेट करें।
- कवरलिप्स के रैक को पानी के साथ स्प्रे करने के लिए एक बोतल का उपयोग करें जब तक कि यह कवरलिप्स को सुचारू रूप से न चला जाए। इथेनॉल के साथ दोहराएं।
- रैक को 20 मिनट के लिए 90 डिग्री सेल्सियस पर ओवन में सूखने के लिए रखें। उपयोग से पहले संदूषण को रोकने के लिए कमरे के तापमान पर अंकित कवरलिप्स के रैक को बंद कंटेनरों में स्टोर करें।
2. एक्टिन फिलामेंट पोलीमराइजेशन
- समाधान T बनाएँ
- 50 एमएल शंक्वाकार ट्यूब में, 3.94 ग्राम ट्राइस-एचसीएल और 0.147 ग्राम सीएसीएल2 जोड़ें। 50 एमएल की कुल मात्रा बनाने के लिए आरओ पानी जोड़ें और अच्छी तरह मिलाएं।
नोट: समाधान टी की अंतिम सांद्रता क्रमशः 500 mM Tris-HCl और 20 mM CaCl2 है। - ट्यूब समाधान टी को लेबल करें और इसे 4 डिग्री सेल्सियस पर स्टोर करें।
- 50 एमएल शंक्वाकार ट्यूब में, 3.94 ग्राम ट्राइस-एचसीएल और 0.147 ग्राम सीएसीएल2 जोड़ें। 50 एमएल की कुल मात्रा बनाने के लिए आरओ पानी जोड़ें और अच्छी तरह मिलाएं।
- टीसी बफर बनाएं
- 50 एमएल शंक्वाकार ट्यूब में 40 एमएल आरओ पानी और 1.5 एमएल सॉल्यूशन टी मिलाएं। केंद्रित कोह की छोटी मात्रा जोड़कर पीएच को 8.0 में बदलें। समाधान के 50 एमएल बनाने के लिए पानी जोड़ें, और पीएच सत्यापित करें। यदि आवश्यक हो तो पीएच समायोजित करें।
नोट: अंतिम टीसी बफर में पीएच 8 पर 5 एमएम ट्राइस-एचसीएल और0.2 एमएम सीएसीएल 2 शामिल हैं। - ट्यूब टीसी को लेबल करें और इसे 4 डिग्री सेल्सियस पर स्टोर करें।
- 50 एमएल शंक्वाकार ट्यूब में 40 एमएल आरओ पानी और 1.5 एमएल सॉल्यूशन टी मिलाएं। केंद्रित कोह की छोटी मात्रा जोड़कर पीएच को 8.0 में बदलें। समाधान के 50 एमएल बनाने के लिए पानी जोड़ें, और पीएच सत्यापित करें। यदि आवश्यक हो तो पीएच समायोजित करें।
- एफसी बफर बनाएं
- 100 एमएल बफर बोतल में 85 एमएल आरओ पानी, 10 एमएल सॉल्यूशन टी, 3.73 ग्राम केसीएल, और 0.041 ग्राम एमजीसीएल2 जोड़ें। केंद्रित KOH की छोटी मात्रा जोड़कर pH को 7.5 में संशोधित करें। 100 एमएल की अंतिम मात्रा बनाने के लिए पानी जोड़ें और पीएच सत्यापित करें।
नोट: अंतिम एफसी बफर में पीएच 7.5 पर 500 एमएम ट्राइस-एचसीएल, 500 एमएम केसीएल, 2 एमएम एमजीसीएल2 और 2 एमएम सीएसीएल2 शामिल हैं। - ट्यूब एफसी को लेबल करें और इसे 4 डिग्री सेल्सियस पर स्टोर करें।
- 100 एमएल बफर बोतल में 85 एमएल आरओ पानी, 10 एमएल सॉल्यूशन टी, 3.73 ग्राम केसीएल, और 0.041 ग्राम एमजीसीएल2 जोड़ें। केंद्रित KOH की छोटी मात्रा जोड़कर pH को 7.5 में संशोधित करें। 100 एमएल की अंतिम मात्रा बनाने के लिए पानी जोड़ें और पीएच सत्यापित करें।
- जनरल एक्टिन बफर (जीएबी) तैयार करें।
- एक माइक्रोसेंट्रीफ्यूज ट्यूब में टीसी बफर के 485 μL, 10 mM ATP के 10 μL और 50 mM DTT के 5 μL मिलाएं।
नोट: अंतिम बफर स्थितियां 5 mM Tris-HCl, 0.2 mM CaCl2, 0.5 mM DTT, और 0.2 mM ATP हैं। - इसे जीएबी के रूप में लेबल करें और इसे 4 डिग्री सेल्सियस पर स्टोर करें।
- एक माइक्रोसेंट्रीफ्यूज ट्यूब में टीसी बफर के 485 μL, 10 mM ATP के 10 μL और 50 mM DTT के 5 μL मिलाएं।
- एक्टिन पोलीमराइजेशन बफर (एपीबी) तैयार करें।
- एक माइक्रोसेंट्रीफ्यूज ट्यूब में एफसी बफर के 455 μL, 100 mM ATP के 25 μL और 50 mM DTT के 20 μL मिलाएं।
नोट: अंतिम बफर स्थितियां 50 mM Tris-HCl, 500 mM KCl, 2 mM MgCl2, 2 mM CaCl2 2 mM DTT, और 5 mM ATP हैं। - ट्यूब को एपीबी के रूप में लेबल करें और इसे 4 डिग्री सेल्सियस पर स्टोर करें।
- एक माइक्रोसेंट्रीफ्यूज ट्यूब में एफसी बफर के 455 μL, 100 mM ATP के 25 μL और 50 mM DTT के 20 μL मिलाएं।
- Reconstitute actin
- लियोफिलाइज्ड एक्टिन की 1 मिलीग्राम शीशी में 100 μL विआयनीकृत पानी जोड़कर खरगोश कंकाल की मांसपेशी एक्टिन का पुनर्गठन करें। धीरे-धीरे ऊपर और नीचे करके अच्छी तरह मिलाएं। 5 μL नमूनों में एलिकोट करें, स्नैप-फ्रीज करें, और 10 मिलीग्राम / एमएल एक्टिन एलिकोट को -80 डिग्री सेल्सियस पर स्टोर करें।
- आरओ पानी के 20 μL जोड़कर बायोटिनाइलेटेड खरगोश कंकाल की मांसपेशी एक्टिन का पुनर्गठन करें। 5 μL नमूनों में एलिकोट करें, स्नैप-फ्रीज करें, और -80 डिग्री सेल्सियस पर 1 मिलीग्राम / एमएल बायोटिनिलेटेड एक्टिन एलिकोट स्टोर करें।
- रोडामाइन फेलोइडिन स्थिरीकरण के साथ गैर-लेबल एक्टिन पोलीमराइजेशन
- 10 मिलीग्राम/एमएल एक्टिन की एक शीशी को पिघलाकर बर्फ पर रखें।
- ताजा जीएबी बफर तैयार करें, एक्टिन एलिकोट में 100 μL GAB जोड़ें, और धीरे-धीरे ऊपर और नीचे पाइप करके मिलाएं। 1 घंटे के लिए बर्फ पर घोल को इनक्यूबेट करें।
- इनक्यूबेशन के दौरान ताजा एपीबी तैयार करें। इनक्यूबेशन के बाद, एक्टिन समाधान में 11 μL APB जोड़कर एक्टिन को फिलामेंट्स में पॉलीमराइज़ करें। धीरे-धीरे ऊपर और नीचे करके अच्छी तरह मिलाएं। 20 मिनट के लिए बर्फ पर रखें।
- ताजा बहुलककृत एक्टिन फिलामेंट समाधान में रोडामाइन-लेबल फेलोइडिन के 5 μL जोड़ें। 1 घंटे के लिए अंधेरे में बर्फ पर छोड़ दें।
- 4 डिग्री सेल्सियस पर अंधेरे में एल्यूमीनियम पन्नी में लपेटे गए रोडामाइन एक्टिन शीशी स्टोर करें।
नोट: इन फिलामेंट्स का उपयोग अधिकतम 1 सप्ताह की अवधि के लिए करने का सुझाव दिया जाता है। एएफ गुणवत्ता की पुष्टि प्रत्येक दिन एक प्रवाह सेल की त्वरित इमेजिंग के माध्यम से की जा सकती है जिसमें केवल एएफ होते हैं और दिन-प्रतिदिन लगातार फिलामेंट्स देखते हैं।
- एलेक्सा फ्लोर 488 फेलोइडिन स्थिरीकरण के साथ बायोटिनाइलेटेड एक्टिन पोलीमराइजेशन
- 10 मिलीग्राम/एमएल एक्टिन की एक शीशी और 1 मिलीग्राम/एमएल बायोटिनिलेटेड एक्टिन की 1 शीशी को पिघलाकर बर्फ पर रखें।
- ताजा जीएबी बफर बनाएं।
- दो शीशियों (चरण 2.8.1) को 10: 1 एक्टिन: बायोटिनिलेटेड एक्टिन अनुपात में मिलाएं। एक्टिन मिश्रण में 100 μL GAB जोड़ें और धीरे-धीरे ऊपर और नीचे पाइप करके अच्छी तरह मिलाएं। 1 घंटे के लिए बर्फ पर इनक्यूबेट करें।
- इनक्यूबेशन के दौरान ताजा एपीबी बनाएं।
- इनक्यूबेशन चरण के बाद, एक्टिन समाधान में एपीबी के 11 μL जोड़कर एक्टिन को पॉलीमराइज़ करें। धीरे-धीरे ऊपर और नीचे करके अच्छी तरह मिलाएं। 20 मिनट के लिए बर्फ पर इनक्यूबेट करें।
- एलेक्सा फ्लूर 488-लेबल फेलोइडिन का 5 μL जोड़ें और 1 घंटे के लिए अंधेरे में बर्फ पर इनक्यूबेट करें।
- एल्यूमीनियम पन्नी में लपेटी गई बायोटिनिलेटेड एक्टिन शीशी को अंधेरे में 4 डिग्री सेल्सियस पर स्टोर करें।
नोट: इन फिलामेंट्स का उपयोग अधिकतम 1 सप्ताह की अवधि के लिए किया जा सकता है।
3. मायोसिन और मोती की तैयारी
- मायोसिन II का पुनर्गठन करें
- एक मानक मिनीसेंट्रीफ्यूज का उपयोग करके ट्यूब के तल पर एकत्र करने के लिए संक्षेप में (~ 5 एस) लियोफिलाइज्ड कंकाल मायोसिन II को स्पिन करें।
- आरओ पानी में तैयार 1 एमएम डीटीटी के 100 μL जोड़कर मायोसिन को 10 मिलीग्राम / एमएल तक पुनर्गठित करें।
- आरओ पानी में 10 मिलीग्राम / एमएल मायोसिन के 10 μL को 1 mM DTT के 90 μL में जोड़कर स्टॉक मायोसिन समाधान 10x पतला करें। छोटी मात्रा (1-5 μL) एलिकोट बनाएं, स्नैप-फ्रीज करें, और -80 डिग्री सेल्सियस पर स्टोर करें।
नोट: मायोसिन गतिविधि की पुष्टि एक मानक ग्लाइडिंग फिलामेंट परख करके की जा सकती है जैसा कि पहले 46,47 प्रकाशित हुआ था। संक्षिप्त विवरण के लिए चर्चा देखें.
- स्ट्रेप्टाविडिन-लेपित मोतियों की सफाई
- 1 μm स्ट्रेप्टाविडिन मोतियों के 20 μL को 80 μL RO पानी में पतला करें। 9,600 × ग्राम पर घूमकर और आरओ पानी के 100 μL में पुनर्गठित करके चार बार धोएं।
- 40% आयाम पर 2 मिनट के लिए सोनिकेट करें और धोए गए मोतियों को 4 डिग्री सेल्सियस पर एक घूर्णन पर स्टोर करें।
4. प्रवाह सेल तैयारी
- 50 एमएल ट्यूब में 100% इथेनॉल के 30 एमएल जोड़कर और पानी में 0.1% डब्ल्यू /वी पॉली-एल-लाइसिन के 200 μL जोड़कर पॉली-एल-लाइसिन समाधान (पीएलएल) तैयार करें और अच्छी तरह से मिलाएं।
- पीएलएल घोल में एक अंकित कवरस्लिप जोड़ें और इसे 15 मिनट के लिए भिगोने दें। कवरस्लिप को चिमटी से हटा दें, केवल कवरस्लिप के किनारे को छूने का ध्यान रखें क्योंकि इसे ट्यूब से ऊपर खींचा जाता है (चित्रा 1 ए-सी देखें)। एक प्यारे हाथ से उनके किनारों से कवरलिप्स को पकड़ो।
- कवरस्लिप को फ़िल्टर की गई एयरलाइन के साथ तब तक सुखाएं जब तक कि कोई इथेनॉल न बचा हो और कवरस्लिप पर कोई अवशेष न हो।
- एक माइक्रोस्कोप स्लाइड के बीच में डबल-साइडेड चिपचिपा टेप के दो टुकड़े लागू करें, एक दूसरे से 3-4 मिमी अलग। स्लाइड के किनारे से लटकने वाले अतिरिक्त टेप को फाड़ दें या काट दें।
- चैनल बनाने के लिए माइक्रोस्कोप स्लाइड (टी बनाने) की लंबी धुरी के लंबवत टेप के शीर्ष पर पीएलएल-लेपित कवरस्लिप जोड़ें।
- टेप पर कवरस्लिप को संपीड़ित करने के लिए एक छोटी ट्यूब का उपयोग करें और टेप के पारदर्शी होने तक माइक्रोस्कोप स्लाइड को अच्छी तरह से स्लाइड करें (चित्रा 1 ए)। सुनिश्चित करें कि टेप में कोई बुलबुले नहीं हैं क्योंकि इससे प्रवाह चैनल से रिसाव हो सकता है।
नोट: प्रवाह सेल 10-15 μL की मात्रा रख सकता है।
5. एक्टोमायोसिन बंडल तैयारी
- अलग-अलग ट्यूबों में, प्रत्येक प्रकार के एक्टिन फिलामेंट (रोडामाइन- और बायोटिनाइलेटेड 488-लेबल) 600x को संबंधित के 0.5 μL को मिलाकर पतला करें, जिसे एक्टिन को 300 μL APB के साथ लेबल किया गया है। प्रत्येक ट्यूब में तदनुसार लेबल वाले फेलोइडिन का एक अतिरिक्त 5 μL जोड़ें और 15 मिनट के लिए अंधेरे में बर्फ पर इनक्यूबेट करें।
- बायोटिनिलेटेड एक्टिन समाधान में, 500 मिलीग्राम / एमएल पर बीटा-डी-ग्लूकोज के 1 μL की ऑक्सीजन स्कैवेंजिंग प्रणाली जोड़ें, 25 मिलीग्राम / एमएल पर ग्लूकोज ऑक्सीडेज का 1 μL, और 500 इकाइयों / एमएल पर कैटालेज का 1 μL जोड़ें। 100 mM ATP का 1 μL और 100x पतला, साफ किए गए स्ट्रेप्टाविडिन मोती का 1 μL जोड़ें। धीरे से पिपेट टिप के साथ हिलाएं। निलंबन को 4 डिग्री सेल्सियस पर एक घूर्णन पर रखें, जबकि बाकी एक्टोमायोसिन बंडल को इकट्ठा किया जा रहा है।
- पीएलएल प्रवाह सेल (चित्रा 1 डी) में पतला रोडामाइन एक्टिन का 15 μL जोड़ें। प्रवाह सेल के माध्यम से अतिरिक्त समाधान को विक करें लेकिन प्रवाह चैनल को सूखने न दें। आर्द्रता कक्ष में 10 मिनट के लिए इनक्यूबेट करें।
नोट: आर्द्रता कक्षों को खाली पिपेट टिप बक्से से बनाया जा सकता है, जिसमें नीचे पानी जोड़ा जाता है और प्रकाश को अवरुद्ध करने के लिए एल्यूमीनियम पन्नी में ढक्कन कवर किया जाता है। - एपीबी में 1 मिलीग्राम / एमएल कैसिइन समाधान तैयार करें।
- बाद के घटकों के गैर-विशिष्ट बंधन को रोकने के लिए 1 मिलीग्राम / एमएल कैसिइन का 15 μL जोड़ें (चित्रा 1 ई)। आर्द्रता कक्ष में 5 मिनट के लिए इनक्यूबेट करें।
- चरण 5.2 से बायोटिनिलेटेड एक्टिन और बीड सस्पेंशन में मायोसिन की वांछित एकाग्रता जोड़ें। धीरे से पिपेट टिप के साथ हिलाएं, और फिर तुरंत चरण 5.2 निलंबन के 15 μL + प्रवाह सेल में वांछित मायोसिन एकाग्रता जोड़ें (चित्रा 1 एफ, जी)। 20 मिनट के लिए इनक्यूबेट करें। इमेजिंग और ऑप्टिकल ट्रैपिंग प्रयोगों के दौरान वाष्पीकरण को रोकने के लिए नेल पॉलिश के साथ प्रवाह सेल के खुले सिरों को सील करें।
नोट: 1 μM की मायोसिन समाधान एकाग्रता मजबूत बंडलिंग पैदा करती है और परख के वांछित अनुकूलन के लिए एक प्रारंभिक बिंदु के रूप में इस्तेमाल किया जा सकता है ( चित्रा 2 देखें)।
6. ऑप्टिकल ट्रैप (एनटी 2 नैनोट्रैकर 2) का उपयोग कर बल माप
नोट: जबकि नीचे दिया गया प्रोटोकॉल विशेष रूप से एनटी 2 सिस्टम के लिए है, इस परख का उपयोग अन्य ऑप्टिकल ट्रैपिंग सिस्टम के साथ किया जा सकता है, जिसमें कस्टम-निर्मित भी शामिल हैं, जिनमें प्रतिदीप्ति क्षमताएं भी हैं। सामान्य वर्कफ़्लो फोकस में स्लाइड की सतह प्राप्त करने, मोती अंशांकन करने और फ्लोरोसेंट एक्टिन बंडलों को खोजकर डेटा प्राप्त करने के समान रहता है। एनटी 2 प्रणाली के लिए, पूरक चित्रा एस 1, पूरक चित्रा एस 2, पूरक चित्रा एस 3, पूरक चित्रा एस 4, पूरक चित्रा एस 5, पूरक चित्रा एस 6, और पूरक चित्रा एस 7 ऑप्टिकल ट्रैपिंग सिस्टम और सॉफ्टवेयर इंटरफ़ेस का विवरण प्रदान करते हैं।
- नियंत्रण बॉक्स और लेजर चालू करें (पूरक चित्रा एस 1)।
- डेस्कटॉप पर जेपीके नैनोट्रैकर आइकन पर क्लिक करके ऑप्टिकल ट्रैप कंप्यूटर सॉफ्टवेयर प्रारंभ करें।
- केंद्र में Logitech बटन पर क्लिक करके दूरस्थ नियंत्रक को जगाएं (पूरक चित्रा S2)।
- ऑन/ऑफ स्विच (पूरक चित्रा एस 3) को घुमाकर फ्लोरेसेंस मॉड्यूल चालू करें।
- ब्राइटफील्ड इमेजिंग (पूरक चित्रा एस 4) के लिए फ़िल्टर क्यूब बुर्ज चालू करें।
- एक बार सिस्टम तैयार हो जाने के बाद, स्क्रीन के बाएं-निचले कोने पर लेजर पावर बटन का उपयोग करके लेजर को 50 मेगावाट तक चालू करें और इसे 30 मिनट (पूरक चित्रा एस 5) के लिए स्थिर होने दें।
- प्रयोग के दौरान देखने और हेरफेर के लिए उन खिड़कियों को लाने के लिए सॉफ्टवेयर के भीतर रोशनी, कैमरा, उद्देश्य और स्टेज मूवमेंट बटन पर क्रमिक रूप से क्लिक करें। ऑन /ऑफ बटन पर क्लिक करके माइक्रोस्कोप रोशनी को चालू करें और बार को दाईं ओर क्लिक करके और खींचकर इसे अधिकतम शक्ति पर सेट करें (पूरक चित्रा एस 5)।
- नमूना क्षेत्र खोलें और माइक्रोस्कोप चरण से नमूना धारक को हटा दें। प्रवाह सेल जोड़ें, इसे धातु नमूना धारकों के साथ सुरक्षित करें, और सुनिश्चित करें कि कवरस्लिप के साथ स्लाइड नीचे है।
- निचले उद्देश्य के केंद्र में 30 μL RO पानी जोड़ें। पिपेट टिप को लेंस को छूने न दें। नमूना चरण को पुन: सम्मिलित करें।
नोट: चूंकि एनटी 2 सिस्टम ट्रैपिंग उद्देश्य के रूप में एक जल विसर्जन उद्देश्य का उपयोग करता है, इसलिए उपयोगकर्ता के सेटअप में ट्रैपिंग उद्देश्य के आधार पर विसर्जन मीडिया अलग-अलग हो सकता है। - रिमोट कंट्रोलर पर ऑन-स्क्रीन कंट्रोल तीर या एल 2 का उपयोग करके निचले उद्देश्य को उठाएं जब तक कि पानी का मोती कवरस्लिप (पूरक चित्रा एस 5) को नहीं छूता है।
- रिमोट कंट्रोलर पर ऑन-स्क्रीन तीर या आर 2 का उपयोग करके प्रवाह सेल की लगभग आधी दूरी तक पहुंचने तक शीर्ष उद्देश्य को कम करें। शीर्ष उद्देश्य के तहत सीधे प्रवाह सेल के शीर्ष पर 170 μL आरओ पानी जोड़ें। शीर्ष उद्देश्य को तब तक कम करें जब तक कि यह पानी की सतह के तनाव को तोड़ न दे और एक मेनिस्कस न बना दे।
- रिमोट कंट्रोलर पर तीर पैड का उपयोग करके माइक्रोस्कोप चरण को तब तक ले जाएं जब तक कि प्रवाह चैनल से सटे टेप के किनारे तक नहीं पहुंच जाता। नमूना दरवाजा बंद करें।
नोट: नमूना दरवाजा बंद करने पर एक "क्लिक" इंगित करता है कि लेजर शटर अब खुला है। यह एक सुरक्षा सुविधा है जो केवल दरवाजा बंद होने पर शटर खोलने की अनुमति देती है। - स्क्रीन में ऑब्जेक्टिव विंडो का उपयोग करके, ऑन-स्क्रीन नियंत्रणों का उपयोग करके ऊपरी तीर पर क्लिक करके लेजर ऑब्जेक्टिव नामक निचले उद्देश्य को ऊपर लाकर टेप के किनारे को फोकस में लाएं। नीचे तीर (पूरक चित्रा एस 5) पर क्लिक करके शीर्ष उद्देश्य के लिए भी ऐसा ही करें।
नोट: दोहरे तीर उद्देश्य या चरण को तेजी से आगे बढ़ाते हैं। टेप के किनारे का उपयोग ध्यान केंद्रित करने के लिए किया जाता है क्योंकि यह एक बड़ी, आसानी से मिलने वाली वस्तु है जो कवरस्लिप सतह के करीब है। टेप के भीतर हवा के बुलबुले एक और विकल्प हैं। हालांकि, इसकी आवश्यकता नहीं है यदि उपयोगकर्ता के पास सतह फोकस या पसंदीदा इन-हाउस विधि खोजने के लिए एक स्वचालित दिनचर्या है। - एक बार टेप फोकस में होने के बाद, ऑप्टिकल ट्रैप के शीर्ष पर आईरिस को आंशिक रूप से बंद करें। शीर्ष उद्देश्य को तब तक नीचे लाएं जब तक कि आईरिस का बहुभुज आकार दिखाई न दे। उन किनारों को फोकस में लाएं, आईरिस को फिर से खोलें, और फिर पैडलॉक आइकन (पूरक चित्रा एस 5) पर क्लिक करके उद्देश्यों को जोड़ेँ।
- एक फ्लोटिंग बीड ढूंढें और ट्रैप शटर बटन पर क्लिक करके इसे फंसाएं, जो शटर खोल देगा और ट्रैपिंग लेजर को नमूने को हिट करने की अनुमति देगा। स्क्रीन पर ट्रैप कर्सर पर क्लिक करें और ट्रैपिंग लेजर के स्थान को स्थानांतरित करने के लिए इसे खींचें। एक बार फंसने के बाद, बल और विस्थापन के लिए वोल्टेज माप को सहसंबंधित करने के लिए मोती को कैलिब्रेट करें।
- अंशांकन बटन पर क्लिक करें। पावर स्पेक्ट्रा विश्लेषण के आधार पर अंशांकन दिनचर्या को समायोजित करें और एक्स, वाई और जेड दिशाओं (पूरक चित्रा एस 6) के लिए सॉफ्टवेयर के भीतर कोने की आवृत्ति फिट करें।
- सेटिंग्स पर क्लिक करें। मोती (1,000 एनएम) के व्यास में टाइप करें, और सॉफ्टवेयर विंडो के निचले बाईं ओर पाए गए चरण के तापमान में टाइप करें। (पूरक चित्रा एस 6 देखें)।
- ट्रैप 1 पर क्लिक करें। एक्स सिग्नल पर क्लिक करें। कोने की आवृत्ति फिट करने के लिए रन पर क्लिक करें। फ़ंक्शन फिट को ऑप्टिमाइज़ करने के लिए विंडो के भीतर क्लिक करें और खींचें। संवेदनशीलता और कठोरता मूल्यों के लिए इसका उपयोग करें पर क्लिक करें। मान स्वीकार करें पर क्लिक करें. Y और Z संकेतों के लिए दोहराएं। विंडो बंद करें। (पूरक चित्रा एस 6 देखें)।
नोट: अन्य ऑप्टिकल ट्रैपिंग सिस्टम या कस्टम-निर्मित सिस्टम पर बीड कैलिब्रेशन रूटीन जिन्हें उपयोगकर्ता द्वारा मजबूती से परीक्षण किया गया है, जैसे कि समविभाजन विधि या ड्रैग फोर्स विधि, भी स्वीकार्य हैं57,58। - कवरस्लिप की सतह पर एएफ से बंधे मोतियों की खोज करके एक एक्टोमायोसिन बंडल खोजें।
- जब अन्य फ्लोटिंग मोतियों द्वारा अनक्रॉव्ड एक मोती का पता लगाया जाता है, तो बंडल की उपस्थिति को सत्यापित करने के लिए फ्लोरेसेंस इमेजिंग द्वारा इसके आसपास के एएफ का निरीक्षण करें।
- सत्यापित करें कि फ्लोरोसेंट एएफ कोलोकाइज्ड दोनों की तलाश करके एक बंडल मौजूद है। सफेद प्रकाश स्रोत चालू करें और बुर्ज को मोड़कर प्रत्येक एक्टिन फिलामेंट को चित्रित करने के लिए उपयुक्त फ़िल्टर क्यूब का उपयोग करें (क्रमशः एलेक्सा फ्लूर 488 और रोडामाइन उत्तेजना के लिए 488 एनएम और 532 एनएम उत्तेजना फिल्टर क्यूब्स)। पूरक चित्र S4 देखें।
नोट: एकल एएफ की प्रतिदीप्ति तीव्रता को सत्यापित करने के लिए एक नियंत्रण प्रयोग उन बंडलों की पहचान करने में उपयोगी हो सकता है जो एकल 488- और एकल रोडामाइन-लेबल फिलामेंट्स से बने होते हैं, या जो भी फ्लोरोफोरे का उपयोग करना चुनते हैं, उस पर लागू होते हैं। - एक बार सत्यापित होने के बाद, ट्रैप शटर बटन पर क्लिक करके बंडल के शीर्ष फिलामेंट से जुड़े मोती को फंसाएं।
- ऑसिलोस्कोप बटन (पूरक चित्रा एस 7) पर क्लिक करके डेटा रिकॉर्ड करने के लिए ऑन-स्क्रीन नियंत्रणों का उपयोग करें। डेटा रिकॉर्ड किए बिना माप की कल्पना करने के लिए, स्टार्ट पर क्लिक करें। सभी डेटा सहेजने के लिए, Autosave पर क्लिक करें. माप रिकॉर्ड करने के लिए, प्रारंभ रिकॉर्ड पर क्लिक करें. ड्रॉप डाउन मेनू एक्स सिग्नल या वाई सिग्नल से चुनकर चुनें कि कौन से डेटा को वास्तविक समय (स्थिति, बल, एक्स-दिशा, वाई-दिशा) में विज़ुअलाइज़ किया जाना है। याद रखें कि xdirection बाएं से दाएं है, और वाई-दिशा स्क्रीन पर ऊपर और नीचे है। पूरक चित्र S7 देखें।
नोट: डेटा .out फ़ाइलों के रूप में सहेजा जाएगा और इसमें प्रत्येक दिशा के लिए समय, वोल्टेज, विस्थापन और बल शामिल है। इन फ़ाइलों को विज़ुअलाइज़ेशन और विश्लेषण के लिए अन्य सॉफ़्टवेयर में निर्यात किया जा सकता है।
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
एक्टोमायोसिन बंडल सिस्टम युक्त प्रवाह कोशिकाएं एक मानक डिजाइन की होती हैं, जिसमें एक माइक्रोस्कोप स्लाइड और एक उकेरा हुआ कवरस्लिप होता है जो दो तरफा चिपचिपा टेप (चित्रा 1) से बने चैनल द्वारा अलग किया जाता है। परख को तब प्रोटोकॉल में वर्णित मंचित परिचय का उपयोग करके कवरस्लिप से बनाया जाता है। अंतिम परख में टेम्पलेट रोडामाइन-लेबल एक्टिन फिलामेंट्स होते हैं; वांछित मायोसिन एकाग्रता ( चित्रा 2 और चित्रा 3 में प्रतिनिधि परिणामों के लिए 1 μM का उपयोग किया गया था); बायोटिनाइलेटेड, एलेक्सा फ्लोर 488-लेबल एक्टिन फिलामेंट्स; 1 चम्मच मोती; ऑक्सीजन स्कैवेंजिंग सिस्टम; एटीपी; और एपीबी बफर। प्रति प्रवाह सेल में कई बंडल बनेंगे, और ऊपर वर्णित एक्टिन सांद्रता बंडलों के बीच पर्याप्त अंतर देती है ताकि यह सुनिश्चित हो सके कि कोई अवांछित इंटरैक्शन न हो। यह डेटा अधिग्रहण दक्षता बढ़ाने के लिए प्रति प्रवाह सेल कई बल माप प्राप्त करने की सुविधा भी प्रदान करता है। बल प्रोफाइल एक प्रवाह सेल के भीतर और प्रवाह सेल से प्रवाह सेल तक प्रतिलिपि प्रस्तुत करने योग्य होना चाहिए।
जबकि उपरोक्त प्रोटोकॉल एक वाणिज्यिक ऑप्टिकल ट्रैपिंग सेटअप के उपयोग की ओर अग्रसर है, यहां प्रस्तुत प्रवाह सेल और परख का उपयोग माइक्रोस्कोप या माइक्रोस्कोप चरण के साथ युग्मित एक अलग वाणिज्यिक उपकरण या कस्टम-निर्मित ऑप्टिकल ट्रैपिंग सेटअप के लिए आसानी से किया जा सकता है और प्रतिदीप्ति इमेजिंग क्षमताएं हैं। एक बार उपरोक्त प्रोटोकॉल के अनुसार सभी प्रवाह सेल परिवर्धन पूरा हो जाने के बाद, स्लाइड (चित्रा 1) पर एक्टोमायोसिन बंडल तत्काल माप के लिए तैयार होते हैं। प्रवाह सेल को ऑप्टिकल ट्रैप माइक्रोस्कोप चरण में जोड़ा जाता है, कई मोती अंशांकन माप प्राप्त किए जाते हैं, और बंडल फिलामेंट्स के फ्लोरेसेंस कोलोकलाइजेशन के माध्यम से बंडलों की पहचान की जाती है। एक बंडल से बंधा एक मोती फंस जाता है, और विस्थापन और संबंधित बल माप शुरू होता है। उपयोगकर्ता कंप्यूटर मॉनिटर पर वास्तविक समय में डेटा के अधिग्रहण का निरीक्षण कर सकता है। प्रवाह सेल में उपयोग किए जाने वाले मायोसिन की एकाग्रता के आधार पर, बंडल तुरंत पर्याप्त आंदोलन का प्रदर्शन करना शुरू कर सकता है, या विस्थापन / बल में वृद्धि को प्रभावी ढंग से देखने में 30 एस -1 मिनट लग सकता है।
एक प्रतिनिधि बल ट्रेस चित्रा 3 ए में दिखाया गया है जहां मायोसिन मोटर्स एक पठार के बाद बल में एक स्थिर रैंप प्रदर्शित करते हैं। इस प्रकार के निशान 2-5 मिनट में विकसित होते देखना विशिष्ट है। हालांकि, एक्टोमायोसिन बंडलों को मापना भी संभव है जो कोई शुद्ध बल उत्पन्न नहीं करते हैं (चित्रा 3 बी)। ये निशान आधारभूत शोर के रूप में दिखाई देते हैं या 90 सेकंड से अधिक बल में कोई पर्याप्त शुद्ध वृद्धि नहीं दिखाते हैं। यह मोटर की कम स्थानीय एकाग्रता के कारण होने की संभावना है जो उत्पादक स्लाइडिंग की अनुमति नहीं देता है, या बंडल एक प्रतिकूल समानांतर अभिविन्यास में है जहां फिलामेंट्स के प्लस और माइनस सिरों को संरेखित किया जाता है।
चूंकि प्रवाह सेल की सामग्री घटना रोशनी और ट्रैपिंग लेजर, समय के साथ स्लाइड पर स्थानीय हीटिंग और कट्टरपंथी ऑक्सीजन प्रजातियों की पीढ़ी से गिरावट के लिए अतिसंवेदनशील हो सकती है, इसलिए 1 घंटे से अधिक समय तक एक ही प्रवाह सेल का उपयोग न करने की दृढ़ता से सलाह दी जाती है। अधिकतम दक्षता के लिए, डेटा प्राप्त करते समय एक और परख इनक्यूबेशन करने का सुझाव दिया जाता है। फोर्स ट्रेस को ऑप्टिकल ट्रैपिंग सॉफ्टवेयर से एक्सेल, मैटलैब, इगोर, या अन्य डेटा प्रबंधन कार्यक्रमों में आगे फ़िल्टरिंग और विश्लेषण के लिए निर्यात किया जा सकता है। बंडल प्रयोगों से निकाले जा सकने वाले डेटा में अलग-अलग परख स्थितियों के तहत विभिन्न प्रकार के बल उत्पादन प्रोफाइल (बेसलाइन, रैंप / पठार), बल उत्पादन का वेग, अधिकतम बल उत्पादन, चरण आकार के माध्यम से एन्सेंबल काइनेटिक और स्टेपिंग व्यवहार और चरणों की टीमों के बीच समय, साथ ही ड्यूटी अनुपात शामिल हैं। उपयोगकर्ता परख स्थितियों को यह तुलना करने के लिए भी बदल सकता है कि विभिन्न प्रकार के मायोसिन मोटर्स को जोड़ना, एक्टिन बाइंडिंग प्रोटीन जोड़ना, या बफर स्थितियों को बदलना इन पहनावा बल पीढ़ी विशेषताओं को कैसे प्रभावित करता है।
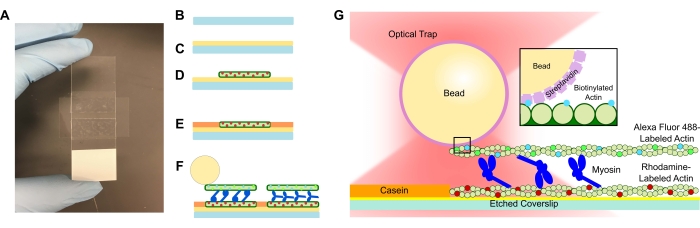
चित्र 1: परख योजनाबद्ध। (ए-सी) अंकित कवरलिप्स को पॉली-एल-लाइसिन में लेपित किया जाता है और डबल-साइडेड टेप और माइक्रोस्कोप स्लाइड का उपयोग करके प्रवाह सेल बनाने के लिए उपयोग किया जाता है। प्रोटोकॉल में वर्णित समयबद्ध परिचय और इनक्यूबेशन चरणों के परिणामस्वरूप रोडामाइन-लेबल फेलोइडिन-स्थिर एक्टिन टेम्पलेट या बॉटम फिलामेंट (डी) के रूप में होता है, इसके बाद गैर-विशिष्ट बाइंडिंग (ई) को रोकने के लिए कैसिइन ब्लॉकिंग होती है, और (एफ) एलेक्सा फ्लोर 488 फेलोइडिन-स्टेबलाइज्ड बायोटिनिलेटेड एक्टिन कार्गो या टॉप फिलामेंट के रूप में, और मायोसिन II की टीमें जो फिलामेंट्स को अलग करती हैं और एटीपी पेश किए जाने पर बल उत्पन्न करती हैं। मोटर्स की ज्यामिति और बंडल के भीतर क्रॉसलिंकिंग की प्रकृति विभिन्न परिस्थितियों में भिन्न हो सकती है, जैसे नमक एकाग्रता59। पिछले अध्ययनों से पता चला है कि मायोसिन टेल डोमेन में एक्टिन फिलामेंट्स और धीमी एन्सेंबल गतिशीलता46 के साथ बातचीत करने की क्षमता है। हालांकि, भारी मेरोमायोसिन प्रयोगों में मायोसिन सिर प्रत्येक सिर को आसन्न एक्टिन फिलामेंट्स60 से बांधने का प्रदर्शन करते हैं। (जी) स्ट्रेप्टाविडिन मोतियों का उपयोग जाल के लिए ऑप्टिकल हैंडल के रूप में किया जाता है और पूरी तरह से कार्गो बायोटिनिलेटेड एक्टिन फिलामेंट से जुड़ता है, जो यह सत्यापित करने में सहायता करता है कि स्लाइड पर उचित बंडल बनते हैं। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें।

चित्रा 2: फ्लोरोसेंट एक्टोमायोसिन बंडल। चित्रा 1 में प्रस्तुत बंडल परख के भीतर एक्टिन फिलामेंट्स और बंडलों के चार अलग-अलग मुठभेड़। एलेक्सा फ्लूर 488 फेलोइडिन चैनल के साथ शीर्ष कार्गो बायोटिनिलेटेड एक्टिन फिलामेंट बाईं ओर दिखाया गया है, और रोडामाइन फेलोइडिन चैनल के साथ नीचे टेम्पलेट एक्टिन फिलामेंट दाईं ओर है। नीचे, एक ही आकृति को आंखों का मार्गदर्शन करने में मदद करने के लिए रंगीन रेखाओं के साथ दिखाया गया है। (ए) एक शीर्ष एक्टिन फिलामेंट एक नीचे एक्टिन फिलामेंट के पास पाया जाता है लेकिन इसमें एक अधूरा ओवरले होता है। इसका उपयोग बंडल प्रयोगों के लिए नहीं किया जाएगा। (बी) शीर्ष और निचले एक्टिन फिलामेंट्स को स्थानीयकृत किया जाता है, और प्रत्येक फिलामेंट की तीव्रता पुष्टि करती है कि वे बंडल के भीतर प्रत्येक एकल फिलामेंट्स हैं। यह बंडल प्रयोगों के लिए एक अच्छा उम्मीदवार होगा। (सी) स्व-इकट्ठे रोडामाइन फिलामेंट्स का एक बड़ा बंडल तल पर पाया जाता है। जबकि एक संबंधित शीर्ष एक्टिन फिलामेंट है जो कोलोकाइज्ड है, बहुत सारे नीचे फिलामेंट्स मौजूद हैं; इस प्रकार, इसका उपयोग बंडल प्रयोगों के लिए नहीं किया जाएगा। यह भी एक उदाहरण है कि जब एक ही प्रकार के कई एक्टिन फिलामेंट्स को बंडल किया जाता है, तो प्रतिदीप्ति तीव्रता बढ़ जाती है। उपयोगकर्ता इसे एक ही फिलामेंट प्रकार के एकल फिलामेंट्स बनाम बंडलों को पहचानने के लिए एक गेज के रूप में उपयोग कर सकता है। (डी) एक निचला फिलामेंट मौजूद है जिसमें कोई संबंधित शीर्ष फिलामेंट नहीं है, यह भी पुष्टि करता है कि कोई रक्तस्राव नहीं हुआ है। इसका उपयोग बंडल प्रयोगों के लिए नहीं किया जाएगा। हम ध्यान दें कि एलेक्सा फ्लूर 488 चैनल में फिलामेंट्स की तीव्रता कम है और विश्वास है कि यह फ़िल्टर सेट के कारण है जिसका उपयोग किया जा रहा है (ज़ीस से फ़िल्टर सेट 09)। रोडामाइन चैनल के लिए उपयोग किया जाने वाला फ़िल्टर सेट ज़ीस से फ़िल्टर सेट 43 है। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें।

चित्रा 3: मायोसिन द्वितीय एन्सेंबल फोर्स जनरेशन। निर्मित इन विट्रो एक्टिन संरचनात्मक पदानुक्रम के भीतर कंकाल मायोसिन द्वितीय मोटर्स के प्रतिनिधि निशान बल उत्पन्न करते हैं। मायोसिन मोटर्स सामूहिक रूप से और उत्पादक रूप से बल उत्पन्न करने के लिए एक साथ काम कर रहे हैं जब तक कि एक पठार तक नहीं पहुंच जाता है और बल को बनाए रखा जाता है (ए) या बेसलाइन (बी) के पास विरोध का अनुभव होता है। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें।
पूरक चित्रा एस 1: ब्रुकर / जेपीके नैनोट्रैकर 2 ऑप्टिकल ट्रैप। (ए) कंप्यूटर मॉनिटर। (बी) कंप्यूटर कीबोर्ड। (ग) कंप्यूटर टॉवर। (डी) नियंत्रक बॉक्स। (ई) लेजर बिजली की आपूर्ति। (एफ) ऑप्टिकल ट्रैप ऑप्टिक्स बॉक्स। (जी) उल्टे माइक्रोस्कोप। (एच) माइक्रोस्कोप चरण के लिए दरवाजा। (I) ब्राइटफील्ड और डिफरेंशियल इंटरफेरेंस कंट्रास्ट इमेजिंग के बीच स्विच करने के लिए पोलराइज़र स्लाइडर। कृपया इस फ़ाइल को डाउनलोड करने के लिए यहाँ क्लिक करें.
पूरक चित्रा एस 2: ऑप्टिकल ट्रैप के लिए रिमोट कंट्रोल। (ए) मोटर चालित चरण को स्थिति में रखने के लिए कीपैड। (बी-सी) जाल की स्थिति समायोजित करें. (D) A, X, और B क्रमशः मुख्य शटर को चालू और बंद करते हैं, 1 शटर को फंसाते हैं, और ट्रैप 2 शटर को फंसाते हैं। (ई) नियंत्रक को जगाने के लिए लॉजिटेक बटन का उपयोग किया जाता है। (एफ) ट्रैपिंग उद्देश्य को स्थिति में रखने के लिए उपयोग किए जाने वाले ऊपर और नीचे के बटन। (जी) अप और डाउन बटन जो डिटेक्शन उद्देश्य को स्थिति में रखने के लिए उपयोग किए जाते हैं। ध्यान दें कि रिमोट कंट्रोल की आवश्यकता नहीं है, और इन सभी जोड़तोड़ को सॉफ्टवेयर में पूरा किया जा सकता है। हालांकि, माइक्रोस्कोप चरण के वातावरण में देखते हुए उद्देश्यों और चरण की स्थिति को नियंत्रित करने में सक्षम होना सुविधाजनक है। कृपया इस फ़ाइल को डाउनलोड करने के लिए यहाँ क्लिक करें.
पूरक चित्रा एस 3: ऑप्टिकल ट्रैप के लिए फ्लोरेसेंस मॉड्यूल। 89 नॉर्थ फोटोफ्लोर फ्लोरेसेंस व्हाइट लाइट सोर्स को उल्टे माइक्रोस्कोप के पीछे युग्मित किया जाता है। इसे टॉगल स्विच (तीर) के साथ चालू और बंद किया जाता है। कृपया इस फ़ाइल को डाउनलोड करने के लिए यहाँ क्लिक करें.
पूरक चित्रा एस 4: फ्लोरेसेंस फिल्टर क्यूब बुर्ज। बुर्ज (तीर) को डीआईसी, रोडामाइन, या एलेक्सा फ्लूर 488 रंगों में इमेजिंग के लिए आवश्यक फ़िल्टर क्यूब का उपयोग करने के लिए चालू किया जा सकता है। ध्यान दें कि विभिन्न फ्लोरोफोरे का उपयोग करने के लिए सेटअप को अनुकूलित करने के लिए फ़िल्टर क्यूब्स को स्विच आउट किया जा सकता है। कृपया इस फ़ाइल को डाउनलोड करने के लिए यहाँ क्लिक करें.
पूरक चित्रा एस 5: नैनोट्रैकर 2 सॉफ्टवेयर। (ए) लेजर पावर बटन और नियंत्रण। (बी) ऑब्जेक्टिव पोजिशनिंग विंडो। दिशात्मक तीर का उपयोग पहचान (ऊपर) और ट्रैपिंग (नीचे) उद्देश्यों को स्थानांतरित करने के लिए किया जाता है। दोहरे तीर उद्देश्यों को उच्च गति से आगे बढ़ाते हैं। नीचे-बाईं ओर नीला और लाल बटन उद्देश्यों को अनकपल करता है और उन्हें उनकी मूल स्थिति में वापस ले लेता है। माइक्रोस्कोप चरण के अंदर और बाहर नमूने लेते समय यह आवश्यक है। उद्देश्यों और पैडलॉक आइकन के साथ बाईं ओर से तीसरा बटन उद्देश्यों को "जोड़ों" के साथ जोड़ता है ताकि जब वे दोनों फोकस में हों और कोहलर रोशनी प्राप्त करें, तो उपयोगकर्ता जेड-अक्ष में ट्रैपिंग और डिटेक्शन उद्देश्यों दोनों को ऊपर और नीचे ले जा सके। (सी) नमूना स्थिति विंडो का उपयोग माइक्रोस्कोप चरण को एक्स- और वाई-अक्ष में स्थानांतरित करने के लिए किया जाता है। दोहरे तीर उच्च गति से मंच को आगे बढ़ाते हैं। यह विंडो शीर्ष मेनू पर ऊपर/नीचे और बाएं/दाएं तीर आइकन पर क्लिक करके सक्रिय होती है। (डी) कैमरा विज़ुअलाइज़ेशन विंडो। रिंच आइकन का उपयोग अनुकूलित इमेजिंग स्थितियों को सेट करने के लिए किया जा सकता है। यह विंडो शीर्ष मेनू पर कैमरा आइकन पर क्लिक करके सक्रिय होती है। (ई) माइक्रोस्कोप रोशनी खिड़की। यह विंडो शीर्ष मेनू पर लाइट बल्ब आइकन पर क्लिक करके सक्रिय होती है। कृपया इस फ़ाइल को डाउनलोड करने के लिए यहाँ क्लिक करें.
पूरक चित्रा एस 6: अंशांकन विंडो। (ए) इस विंडो का उपयोग मोती अंशांकन के लिए किया जाता है और शीर्ष मेनू पर कैल आइकन पर क्लिक करके सक्रिय किया जाता है। एक मोती को कैलिब्रेट करने के लिए, एक्स, वाई और जेड सिग्नल में कोने की आवृत्ति का सबसे अच्छा फिट पूरा किया जाता है। (बी) प्रत्येक सिग्नल के लिए, ऊपर बाईं ओर उपयुक्त सिग्नल बटन चुनें। (सी) रन पर क्लिक करें और हरी विंडो (डी) के भीतर क्लिक करके और खींचकर फिट को अनुकूलित करें। (ई) एक बार फिट से संतुष्ट होने के बाद, संवेदनशीलता और कठोरता के लिए इसका उपयोग करें पर क्लिक करें। यह नैनोमीटर में विस्थापन और पिकोनेवटन में बल को रिकॉर्ड करने की अनुमति देगा। (एफ) फिर, नीचे बाईं ओर मान स्वीकार करें पर क्लिक करें। y और z दिशाओं के लिए दोहराएँ। कृपया इस फ़ाइल को डाउनलोड करने के लिए यहाँ क्लिक करें.
पूरक चित्रा एस 7: डेटा अधिग्रहण विंडो। इस विंडो का उपयोग स्थिति प्राप्त करने और डेटा को बल देने के लिए किया जाता है और उपयोगकर्ता को वास्तविक समय में माप देखने की अनुमति देता है। (ए) यह विंडो शीर्ष मेनू पर एक्स, टी आइकन पर क्लिक करके सक्रिय होती है। (बी) उपयोगकर्ता एक्स और वाई सिग्नल देखने के बीच स्विच कर सकता है। (C) डेटा की कल्पना शुरू करने के लिए प्रारंभ पर क्लिक करें। डेटा सहेजने के लिए ऑटोसेव पर क्लिक करें। डेटा रिकॉर्ड करना और सहेजना शुरू करने के लिए प्रारंभ रिकॉर्ड पर क्लिक करें. कृपया इस फ़ाइल को डाउनलोड करने के लिए यहाँ क्लिक करें.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
फ्लोरेसेंस इमेजिंग के साथ संयुक्त ऑप्टिकल ट्वीज़र्स का उपयोग करके एक इन विट्रो अध्ययन एक्टिन फिलामेंट्स के साथ बातचीत करने वाले मायोसिन एन्सेंबल की गतिशीलता की जांच करने के लिए किया गया था। एक्टिन-मायोसिन-एक्टिन बंडलों को मांसपेशी मायोसिन II, बंडल के तल और कवरस्लिप सतह पर रोडामाइन एक्टिन और बंडल के शीर्ष पर 488-लेबल, बायोटिनिलेटेड एक्टिन फिलामेंट्स का उपयोग करके इकट्ठा किया गया था। खरगोश की मांसपेशियों से एक्टिन प्रोटीन को सामान्य एक्टिन बफर (जीएबी) और एक्टिन पॉलीमराइजिंग बफर (एपीबी) का उपयोग करके पॉलीमराइज्ड और स्थिर किया गया था। जीएबी और एपीबी को एटीपी, एफसी बफर और टीसी बफर का उपयोग करके प्रयोगशाला में हर दिन ताजा तैयार किया जाना चाहिए। मांसपेशी मायोसिन II का उपयोग एक्टिन-मायोसिन-एक्टिन सैंडविच बनाने के लिए किया गया था। फेलोइडिन का उपयोग एक्टिन फिलामेंट्स के फ्लोरोसेंट धुंधला होने के साथ-साथ विट्रो में स्थिरीकरण के लिए किया गया था।
मायोसिन गतिविधि की पुष्टि एक मानक ग्लाइडिंग फिलामेंट परख करके की जा सकती है जैसा कि पहले 46,47 प्रकाशित हुआ था। मायोसिन II और इसके उप-टुकड़े विभिन्न प्रकार के झुकावों में कवरस्लिप सतह से जुड़ सकते हैं, और पूंछ डोमेन की उपस्थिति भारी मेरोमायोसिन46,48,49 का उपयोग करके परख की तुलना में फिलामेंट स्लाइडिंग को धीमा कर सकती है। हालांकि, ग्लाइडिंग और सतह आंदोलन अभी भी देखा जा सकता है। मायोसिन गतिविधि का एक और स्पष्ट प्रदर्शन सक्रिय एक्टिन फिलामेंट ब्रेकिंग है जिसे देखा जा सकता है जहां लंबे समय तक एक्टिन फिलामेंट्स छोटे टुकड़ों में टूट जाते हैं जो फिर कई दिशाओं में चले जाते हैं। यह सतह पर सक्रिय मोटर्स की उच्च सांद्रता के कारण होता है, कई प्रयोगशालाओं द्वारा देखा गया है, और सक्रिय मायोसिन मोटर्स के बिना42,50,51,52,53,54 मौजूद नहीं होता है। इसके अलावा, यहां प्रस्तुत बंडल परख गतिशीलता के मुद्दों को कम करने में सहायता करती है जो मुख्य रूप से ग्लाइडिंग फिलामेंट परख से जुड़ी हुई हैं, जैसे कि ग्लास कवरस्लिप पर मोटर बाइंडिंग ओरिएंटेशन की विविधता, क्योंकि बंडल परख में कांच की सतह को अवरुद्ध करना शामिल है ताकि मोटर्स बंडल 47,55,56 के भीतर बंध जाएं।
पहला कदम एक प्रवाह सेल में पॉली-एल-लाइसिन लेपित कवरस्लिप में नीचे या टेम्पलेट फिलामेंट के रूप में रोडामाइन एक्टिन फिलामेंट्स को जोड़ना है। पॉली-एल-लाइसिन का उपयोग एक्टिन बाइंडिंग को बढ़ावा देने के लिए किया जाता है क्योंकि पॉली-लाइसिन को सकारात्मक रूप से चार्ज किया जाता है जबकि एक्टिन में नकारात्मक चार्ज होते हैं और इसका उपयोग पिछले साइटोस्केलेटल इन विट्रो परख तैयारी 61,62,63 में किया गया है। बंडल गठन से पहले, एक्टिन एकाग्रता को अनुकूलित करने के लिए एक प्रवाह सेल में विभिन्न एक्टिन कमजोर पड़ने को जोड़ा गया था। इस मामले में, स्टॉक से 600x इष्टतम कमजोर पड़ने वाला था जिसने बंडल गठन के लिए पर्याप्त संख्या में टेम्पलेट फिलामेंट्स प्राप्त किए, लेकिन पर्याप्त रिक्ति के साथ ताकि बंडलों को व्यक्तिगत किया जा सके। कमजोर पड़ने को एपीबी बफर का उपयोग करके किया गया था। सतह को अवरुद्ध करने और गैर-विशिष्ट बंधन से बचने के लिए रोडामाइन एक्टिन को जोड़ने के बाद कैसिइन की एक परत थी। प्रवाह सेल को 30 मिनट के लिए इनक्यूबेट किया गया था और किसी भी अनबाउंड एक्टिन फिलामेंट्स को धोने के लिए बफर के साथ इनक्यूबेशन के बाद धोया गया था। अंत में, मायोसिन, 488 / बायोटिन एक्टिन, और स्ट्रेप्टाविडिन-लेपित मोतियों का एक संयोजन एक्टिन-मायोसिन बंडल गठन की सुविधा के लिए प्रवाह सेल में जोड़ा गया था। मोती एकाग्रता ऐसी होनी चाहिए कि सतह-बाध्य बंडलों को बांधने के लिए पर्याप्त हो और अंशांकन की सुविधा के लिए निलंबन में पर्याप्त हो। हालांकि, एक मोती एकाग्रता की बहुत अधिक मात्रा लेजर जाल में गिरने और माप को बाधित करने के कारण पड़ोसी मोतियों को फंसाने के प्रयोगों के दौरान कठिनाई पैदा कर सकती है। मायोसिन मोटर्स को स्लाइड में इंजेक्ट करने से ठीक पहले संयोजन में जोड़ा जाता है ताकि मायोसिन मोटर्स कार्गो या शीर्ष बायोटिनिलेटेड एक्टिन फिलामेंट के साथ पूर्वव्यापी रूप से एकत्र न हों और इस प्रकार नीचे रोडामाइन को बायोटिनाइलेटेड एक्टिन फिलामेंट्स को बंडल करने के लिए बांध देंगे।
एनटी 2 ऑप्टिकल ट्रैपिंग सिस्टम संयुक्त ब्राइटफील्ड, डिफरेंशियल इंटरफेरेंस कंट्रास्ट (डीआईसी), और एपिफ्लोरेसेंस इमेजिंग तौर-तरीकों के साथ एक वाणिज्यिक ऑप्टिकल ट्रैप है। यह ज़ीस एक्सियोऑब्जर्वर 3 उल्टे माइक्रोस्कोप के साथ 100x / NA 1.46 और 63x / NA 1.0 जल विसर्जन ट्रैपिंग और पहचान उद्देश्यों के साथ युग्मित है। सिस्टम एक लेजर ट्रैप की क्लिक और ड्रैग ट्रैपिंग क्षमता से लैस है और इसका उपयोग पहले सूचीबद्ध तौर-तरीकों में से किसी में इमेजिंग करते समय किया जा सकता है। फ्लोरेसेंस इमेजिंग का उपयोग करके गठित बंडलों का पता लगाया जाता है और पुष्टि की जाती है। उपयुक्त फिल्टर क्यूब्स (जीएफपी / एफआईटीसी और टीआरआईटीसी / सीवाई 3) के साथ एक सफेद प्रकाश स्रोत होने से फिलामेंट इमेजिंग के बीच तेजी से स्विचिंग की अनुमति मिलती है। ऑप्टिकल चिमटी का उपयोग करके प्रत्येक बल माप लेने से पहले विभिन्न उत्तेजना तरंग दैर्ध्य पर एएफ की कल्पना करके कोलोकलाइज्ड एएफ को सत्यापित किया गया था। चूंकि फिलामेंट्स ऑक्सीजन स्कैवेंजिंग अभिकर्मक के साथ भी जल्दी से फोटोब्लीच कर सकते हैं, इसलिए यह सुझाव दिया जाता है कि शोधकर्ता बंडल प्रयोगों को करने से पहले तीव्रता और एक्सपोजर समय जैसे विज़ुअलाइज़ेशन मापदंडों को अनुकूलित करें।
ऑप्टिकल ट्रैपिंग को बल माप लेने के लिए नियोजित किया गया था, एटीपी की उपस्थिति में स्ट्रेप्टाविडिन मोतियों का उपयोग करके बायोटिनाइलेटेड कार्गो एक्टिन फिलामेंट को बांधने और बल ट्रांसड्यूसर के रूप में मायोसिन बल पीढ़ी को सक्रिय करने के लिए। ऑप्टिकल ट्रैपिंग द्वारा प्राप्त विस्थापन और बल बनाम समय डेटा विश्लेषण के लिए ट्रैपिंग सॉफ्टवेयर से निकाला गया था। हालांकि, वाणिज्यिक ट्रैपिंग सॉफ्टवेयर विश्लेषण दिनचर्या भी प्रदान करता है जिसका उपयोग किया जा सकता है, या अन्य कार्यक्रमों में कस्टम एल्गोरिदम को उपयोगकर्ता द्वारा ट्रैपिंग डेटा की कल्पना और विश्लेषण करने के लिए प्रोग्राम किया जा सकता है। कस्टम ऑप्टिकल ट्रैपिंग सिस्टम पर, उपयोगकर्ता के पास फिल्टर के साथ सफेद प्रकाश स्रोत के बजाय उत्तेजना लेजर हो सकते हैं, जो उपयोग करने के लिए भी स्वीकार्य हैं। इसके अलावा, फ्लोरोसेंट रंगों को मौजूदा उपकरणों के अनुकूल होने के लिए बदला जा सकता है यदि उत्सर्जन स्पेक्ट्रा ओवरलैप नहीं होता है और खून बहने का कारण बनता है।
हम ध्यान दें कि प्रस्तुत परख एक आधारभूत परख है जिसे उपयोगकर्ता द्वारा एक्टोमायोसिन पहनावा यांत्रिकी के दायरे में उनके शोध प्रश्न के आधार पर आगे अनुकूलित किया जा सकता है। सामान्य वर्कफ़्लो को अन्य इन विट्रो साइटोस्केलेटल एन्सेंबल सिस्टम पर भी लागू किया जा सकता है जो रुचि के हो सकते हैं, जैसे कि सूक्ष्मनलिका बंडल परख जो माइटोटिक स्पिंडल 32,61,63,64,65,66 के न्यूनतम मॉडल बनाते हैं। संशोधन ों में शामिल हो सकते हैं लेकिन उपयोगकर्ता के मौजूदा सेटअप के अनुकूल फ्लोरोफोरे लेबल को बदलने तक सीमित नहीं हैं; मायोसिन एकाग्रता, निर्माण, या आइसोटाइप को बदलना; और अन्य पहलुओं के अलावा बफर स्थितियों को टाइट करना।
इस परख का प्रदर्शन करते समय संभावित चुनौतियां संभव हैं। एक्टिन-मायोसिन बंडल बनाते समय, एक्टिन बंडलों के भीतर मायोसिन एकाग्रता स्लाइड में सजातीय नहीं हो सकती है। इसे समायोजित करने के लिए, पूरी स्लाइड में कई बंडलों को मापा जाएगा ताकि यह सुनिश्चित किया जा सके कि मोटर वितरण और बल उत्पादन प्रोफाइल ठीक से नमूना लिया गया है। बल डेटा की व्याख्या के लिए यदि यह आवश्यक है तो बंडल अभिविन्यास को जानना भी चुनौतीपूर्ण है। इस प्रकार, प्रत्येक बंडल के लिए कई परीक्षण लिए जाने चाहिए। ऑप्टिकल ट्रैपिंग हैंडल की तुलना में छोटे आकार के फ्लोरोसेंट जेलसोलिन या जेलसोलिन-लेपित मोतियों के माध्यम से एक्टिन फिलामेंट एंड लेबलिंग भी शामिल हो सकती है। प्रतिदीप्ति इमेजिंग का उपयोग बंडल अभिविन्यास को कम करने के लिए एक्स और वाई घटक बलों को देखने के लिए भी किया जा सकता है। इसके अलावा, चूंकि मायोसिन एकत्रीकरण अवस्था केसीएल के तेजी से कमजोर पड़ने पर होने वाले मोटे फिलामेंट्स के गठन के साथ बफर की आयनिक ताकत से अत्यधिक प्रभावित होती है, बफर नमक एकाग्रता की उचित रूप सेनिगरानी की जानी चाहिए 67,68।
पिछले अध्ययन जो ग्लाइडिंग परख जैसे अन्य इन विट्रो तरीकों का उपयोग करते थे, मायोसिन डोमेन की भूमिका की पहचान करने और मायोसिन और अन्य एक्टिन बाइंडिंग प्रोटीन के बीच विन्यास और बातचीत का अध्ययन करने में सहायक थे। हालांकि, इन विधियों में एक नुकसान है कि मायोसिन को कठोर सतह पर बांधने से मायोसिन मोटर्स के बीच समन्वय की क्षमता सीमित हो जाएगी और इस प्रकार मेकेनोसेंसिंग फीडबैक जो यह निर्धारित करने के लिए होता है कि मोटर पहनावा उच्च या निम्न शुल्क अनुपात मोड 33,35,41,69 में है या नहीं। इसके अलावा, एकल-मायोसिन मोटर नेटवर्क के साथ ऑप्टिकल ट्रैपिंग इस बात की स्पष्ट समझ नहीं देती है कि मायोसिन मोटर्स एक दूसरे के साथ और एक्टिन फिलामेंट्स के साथ कैसे बातचीत करते हैं। यहां विकसित प्रोटोकॉल एक अनुपालन, पदानुक्रमित एक्टिन नेटवर्क के भीतर मायोसिन मोटर पहनावा गतिशीलता की जांच के लिए अनुमति देता है। यह व्यवस्थित जांच की अनुमति देने के लिए अन्य पहलुओं के बीच, एकाग्रता, आइसोफॉर्म और बफर वातावरण जैसे मोटर-फिलामेंट पहनावा विशेषताओं के संदर्भ में भी अनुकूलन योग्य है। प्रस्तुत प्रोटोकॉल अधिक जटिल एक्टोमायोसिन नेटवर्क के भविष्य के अध्ययन के लिए एक मंच है और ऑप्टिकल ट्रैपिंग द्वारा सुगम विस्थापन और बल उत्पादन माप की सटीकता को बनाए रखता है जो पारंपरिक रूप से एकल-अणु अध्ययन के लिए उपयोग किया जाता है।
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
लेखकों के पास घोषित करने के लिए हितों का कोई टकराव नहीं है।
Acknowledgments
यह काम मिसिसिपी ग्रेजुएट स्टूडेंट काउंसिल रिसर्च फैलोशिप (ओए), मिसिसिपी विश्वविद्यालय सैली मैकडॉनेल-बार्क्सडेल ऑनर्स कॉलेज (जेसीडब्ल्यू, जेईआर), अनुदान संख्या एनएनएक्स 15 एएच 78 एच (जेसीडब्ल्यू, डीएनआर) के तहत मिसिसिपी स्पेस ग्रांट कंसोर्टियम और अनुदान संख्या 848586 (डीएनआर) के तहत अमेरिकन हार्ट एसोसिएशन द्वारा समर्थित है।
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
