

Summary
يتم عرض ومناقشة تشكيل حزم أكتوميوزين في المختبر وقياس توليد قوة مجموعة الميوسين باستخدام ملاقط بصرية.
Abstract
الميوسينات هي بروتينات حركية تحلل ATP لتخطو على طول مسارات خيوط الأكتين (AF) وهي ضرورية في العمليات الخلوية مثل الحركة وتقلص العضلات. لفهم آليات توليد القوة الخاصة بهم ، تم التحقيق في الميوسين II على مستوى الجزيء الواحد (SM) وكفرق من المحركات في المختبر باستخدام الطرق الفيزيائية الحيوية مثل الاصطياد البصري.
أظهرت هذه الدراسات أن سلوك توليد قوة الميوسين يمكن أن يختلف اختلافا كبيرا عند الانتقال من مستوى الجزيء الواحد في ترتيب ثلاثي الخرز إلى مجموعات من المحركات التي تعمل معا على خرزة صلبة أو سطح غطاء في ترتيب انزلاقي. ومع ذلك ، فإن إنشاءات الفحص هذه لا تسمح بتقييم ديناميكيات مجموعة الميوسين داخل التسلسل الهرمي الهيكلي اللزج المرن كما تفعل داخل الخلية. لقد طورنا طريقة باستخدام ملاقط بصرية للتحقيق في ميكانيكا توليد القوة بواسطة مجموعات الميوسين التي تتفاعل مع خيوط الأكتين المتعددة.
تسهل حزم الأكتوميوس هذه التحقيق في بيئة هرمية ومتوافقة تلتقط الاتصالات الحركية وإخراج قوة المجموعة. تسمح الطبيعة القابلة للتخصيص للفحص بتغيير الظروف التجريبية لفهم كيف تؤدي التعديلات على مجموعة الميوسين أو حزمة خيوط الأكتين أو البيئة المحيطة إلى مخرجات قوة مختلفة.
Introduction
البروتينات الحركية ضرورية للحياة ، وتحويل الطاقة الكيميائية إلى عمل ميكانيكي1،2،3. تتفاعل محركات الميوسين مع خيوط الأكتين عن طريق اتخاذ خطوات على طول الخيوط المشابهة للمسار ، وتقوم ديناميكيات شبكات الأكتين الميوسين بتنفيذ تقلص العضلات ، وحركية الخلية ، وحلقة الانقباض أثناء الحركية الخلوية ، وحركة البضائع داخل الخلية ، من بين المهام الأساسية الأخرى3،4،5،6،7،8 . نظرا لأن الميوسين له العديد من الأدوار الأساسية ، فإن الفشل في وظائف شبكة الميوسين أكتين يمكن أن يؤدي إلى تطور المرض ، مثل الطفرات في سلسلة الميوسين الثقيلة التي تسبب فرط انقباض القلب في اعتلال عضلة القلب الضخامي (HCM) 9,10,11,12,13,14 . في تقلص العضلات ، تتعاون محركات الميوسين الفردية مع بعضها البعض من خلال العمل كمجموعة لتوفير الطاقة الميكانيكية المطلوبة التي تنفذ الانزلاق النسبي للتركيز البؤري التلقائي4،15،16،17،18. تشكل محركات الميوسين جسورا متقاطعة بين الرجفان الأذيني وتستخدم تغيرات متطابقة بسبب دورتها الميكانيكية الكيميائية للتحرك بشكل جماعي نحو الطرف الشائك للخيوط المحاذاة17،18،19،20،21.
وقد سهل تطوير مقايسات الحركة الكمية في المختبر على مستوى SM باستخدام تقنيات مثل الاصطياد البصري جمع تفاصيل غير مسبوقة عن كيفية عمل محركات الميوسين الفردية ، بما في ذلك قياس توليد قوة SM وأحجام الخطوات 22،23،24،25،26،27،28،29،30 . طور Finer et al. اختبار الاصطياد البصري "ثلاثي الخرز" أو "الدمبل" للتحقيق في ميكانيكا توليد القوة لمحركات myosin II المفردة23,31. نظرا لأن myosin II العضلي يعمل في فرق للتعاقد مع AFs ولكنه غير عملي على مستوى SM ، كان لا بد من إعادة ترتيب اتجاه فحص الاصطياد البصري من نهج الخرزة الكلاسيكية المرتبطة بالمحرك32. لتشكيل فحص الدمبل ، تم استخدام فخاخين بصريتين لتثبيت التركيز البؤري التلقائي فوق محرك الميوسين المرتبط بحبة متصلة بغطاء ، وتم قياس خرج القوة بواسطة المحرك الفردي من خلال حركات التركيز البؤري التلقائي داخل المصيدة23.
ومع ذلك ، فإن قوى SM واستخدام اتجاه فحص محرك واحد / خيط واحد لا يعطي صورة كاملة عن توليد القوة على مستوى النظام لأن العديد من البروتينات الحركية ، بما في ذلك الميوسين الثاني ، لا تعمل بمعزل عن بعضها البعض وغالبا ما لا تعمل كمجموع لأجزائها 15,16,17,32,33,34,35,36 . الهياكل الأكثر تعقيدا التي تشمل أكثر من محرك واحد يتفاعل مع أكثر من خيوط واحدة ضرورية لفهم أفضل للتآزر بين شبكات خيوط الميوسين والأكتين15,32. تم استغلال اتجاه فحص الدمبل للتحقيق في توليد قوة المجموعة الصغيرة من خلال وجود ميوسينات متعددة متصلة بحرزة أو باستخدام خيوط سميكة من الميوسين متصلة بسطح والسماح للمحركات بالتفاعل مع التركيز البؤري التلقائي المعلق 4,23,34,37,38,39,40.
تشمل اختبارات المجموعة الصغيرة الأخرى مقايسة انزلاقية في المختبر حيث يتم طلاء محركات الميوسين على سطح الغطاء ، ويتم استخدام حبة مرتبطة بالتركيز البؤري التلقائي للتحقيق في القوة الناتجة عن فريق المحركات 4,35,36,38,39,40,41,42,43 . في كلتا الحالتين ، ترتبط الميوسينات بسطح صلب - حبة أو غطاء - وتستخدم AF واحدا. في هذه الحالات ، لا تكون المحركات قادرة على التحرك بحرية أو التواصل مع بعضها البعض ، ولا يعكس وجود myosins مقيد بشكل صارم البيئة الهرمية المتوافقة التي ستعمل فيها المحركات معا في ساركومير32. وقد اقترحت دراسات سابقة أن الميوسين II يمكن أن يستشعر بيئته ويتكيف وفقا لذلك مع تغير ظروف التركيز اللزج المرن أو الحركي عن طريق تغيير الخصائص مثل توليد القوة ونسبة الواجب41،44،45. وبالتالي ، هناك حاجة إلى تطوير اختبار الاصطياد البصري الذي يعزز ويلتقط اتصالات المحرك وتوافق النظام لرسم صورة أكثر واقعية للأسس الميكانيكية لتوليد قوة مجموعة الميوسين II.
هنا ، طورنا طريقة لإقران الهيكل الهرمي في المختبر مع الاصطياد البصري عن طريق تشكيل حزم أو سندويشات أكتوميوزين تتكون من محركات ميوسين متعددة تتفاعل بين اثنين من خيوط الأكتين. تتمتع هندسة المقايسة المعيارية هذه بالقدرة على التحقيق المباشر في كيفية تأثير العوامل الجزيئية والبيئية على توليد قوة الميوسين الجماعية. علاوة على ذلك ، فإن التحقيق في آليات توليد القوة من خلال مجموعات الأكتين الميوسين هذه لديه القدرة على المساعدة في نمذجة وفهم كيفية انتشار المهام الخلوية واسعة النطاق ، مثل تقلص العضلات ، من المستوى الجزيئي9،10،13.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. نقش أغطية
- قم بإذابة 100 غرام من KOH في 300 مل من الإيثانول بنسبة 100٪ في كوب 1000 مل. يحرك المزيج بقضيب التحريك حتى تذوب غالبية KOH.
تنبيه: يمكن أن يسبب محلول KOH المركز حروقا وأضرارا للملابس. ارتداء القفازات وحماية العين ومعطف المختبر. - ضع الأغطية بشكل فردي في رفوف تنظيف الغطاء.
ملاحظة: تم تصميم الرفوف بشقوق تحمل أغطية مفردة متباعدة للسماح بالنقش والشطف على كل وجه من جوانب الغطاء ، وفتحات التصريف في القاع ، ومصنوعة من مواد يمكنها تحمل ظروف الحفر القاسية. يمكن أن تكون مصنوعة حسب الطلب أو شراؤها تجاريا. - قم بإعداد وتسمية ثلاثة أكواب سعة 1000 مل: واحدة مع 300 مل من الإيثانول واثنين من الأكواب مع 300 مل من ماء التناضح العكسي (RO).
ملاحظة: هنا ، تم الحصول على مياه RO من جهاز تنقية المياه في المختبر ، ولكن يمكن أيضا شراؤها تجاريا إذا لم يكن جهاز تنقية محلي متاحا. - ضع كل كوب من الأكواب الأربعة في سونيكتور الحمام لتفريغ الغاز لمدة 5 دقائق.
- اغمر رفا من الأغطية في كوب KOH والإيثانول والسونيكات لمدة 5 دقائق.
- انقل رف الأغطية من كوب KOH / الإيثانول إلى الكأس الذي يحتوي على الإيثانول فقط. اغمس الرف لأعلى ولأسفل في الكأس حتى لا يكون هناك خرزة.
ملاحظة: احرص على عدم إزعاج الأغطية أو إسقاط الرف بقوة في الدورق. سيؤدي ذلك إلى خروج الأغطية من الرف أو التسبب في رش كيميائي. - انقل بعناية رف الأغطية من كوب الإيثانول إلى كوب من الماء ، واغمس لأعلى ولأسفل حتى لا يكون هناك خرزة.
- اغمر رف الأغطية في كوب الماء الذي لم يتم استخدامه بعد وقم بسونيك مرة أخرى لمدة 5 دقائق.
- استخدم زجاجة لرش رف الأغطية بالماء حتى ينفد الغطاء بسلاسة. كرر مع الإيثانول.
- ضع الرفوف لتجف في فرن على حرارة 90 درجة مئوية لمدة 20 دقيقة. قم بتخزين رفوف الأغطية المحفورة في درجة حرارة الغرفة في حاويات مغلقة لمنع التلوث قبل الاستخدام.
2. بلمرة خيوط الأكتين
- جعل الحل T
- في أنبوب مخروطي سعة 50 مل ، أضف 3.94 جم من Tris-HCl و 0.147 جم من CaCl2. أضف ماء RO لجعل الحجم الإجمالي 50 مل واخلطه جيدا.
ملاحظة: التركيزات النهائية للمحلول T هي 500 mM Tris-HCl و 20 mM CaCl2 ، على التوالي. - قم بتسمية الأنبوب Solution T وتخزينه عند 4 درجات مئوية.
- في أنبوب مخروطي سعة 50 مل ، أضف 3.94 جم من Tris-HCl و 0.147 جم من CaCl2. أضف ماء RO لجعل الحجم الإجمالي 50 مل واخلطه جيدا.
- جعل TC المخزن المؤقت
- امزج 40 مل من ماء RO و 1.5 مل من المحلول T في أنبوب مخروطي سعة 50 مل. تغيير الرقم الهيدروجيني إلى 8.0 عن طريق إضافة كميات صغيرة من KOH المركزة. أضف الماء لصنع 50 مل من المحلول ، وتحقق من درجة الحموضة. اضبط الرقم الهيدروجيني إذا لزم الأمر.
ملاحظة: يحتوي المخزن المؤقت TC النهائي على 5 mM Tris-HCl و 0.2 mM CaCl2 عند درجة الحموضة 8. - قم بتسمية الأنبوب TC وتخزينه عند 4 درجات مئوية.
- امزج 40 مل من ماء RO و 1.5 مل من المحلول T في أنبوب مخروطي سعة 50 مل. تغيير الرقم الهيدروجيني إلى 8.0 عن طريق إضافة كميات صغيرة من KOH المركزة. أضف الماء لصنع 50 مل من المحلول ، وتحقق من درجة الحموضة. اضبط الرقم الهيدروجيني إذا لزم الأمر.
- اصنع المخزن المؤقت ل FC
- أضف 85 مل من ماء RO و 10 مل من Solution T و 3.73 جم من KCl و 0.041 جم من MgCl2 إلى زجاجة عازلة سعة 100 مل. قم بتعديل الرقم الهيدروجيني إلى 7.5 عن طريق إضافة كميات صغيرة من KOH المركزة. أضف الماء لجعل الحجم النهائي 100 مل وتحقق من الرقم الهيدروجيني.
ملاحظة: يحتوي المخزن المؤقت النهائي ل FC على 500 mM Tris-HCl و 500 mM KCl و 2 mM MgCl 2 و 2 mM CaCl2 عند درجة الحموضة 7.5. - قم بتسمية الأنبوب FC وتخزينه عند 4 درجات مئوية.
- أضف 85 مل من ماء RO و 10 مل من Solution T و 3.73 جم من KCl و 0.041 جم من MgCl2 إلى زجاجة عازلة سعة 100 مل. قم بتعديل الرقم الهيدروجيني إلى 7.5 عن طريق إضافة كميات صغيرة من KOH المركزة. أضف الماء لجعل الحجم النهائي 100 مل وتحقق من الرقم الهيدروجيني.
- إعداد المخزن المؤقت العام للأكتين (GAB).
- امزج 485 ميكرولتر من المخزن المؤقت TC ، و 10 ميكرولتر من 10 mM ATP ، و 5 ميكرولتر من 50 mM DTT في أنبوب جهاز طرد مركزي دقيق.
ملاحظة: شروط التخزين المؤقت النهائية هي 5 mM Tris-HCl و 0.2 mM CaCl 2 و 0.5 mM DTT و0.2 mM ATP. - قم بتسميته على أنه GAB وقم بتخزينه عند 4 درجات مئوية.
- امزج 485 ميكرولتر من المخزن المؤقت TC ، و 10 ميكرولتر من 10 mM ATP ، و 5 ميكرولتر من 50 mM DTT في أنبوب جهاز طرد مركزي دقيق.
- تحضير المخزن المؤقت لبلمرة الأكتين (APB).
- امزج 455 ميكرولتر من المخزن المؤقت FC ، و 25 ميكرولتر من 100 mM ATP ، و 20 ميكرولتر من 50 mM DTT في أنبوب جهاز طرد مركزي دقيق.
ملاحظة: شروط التخزين المؤقت النهائية هي 50 mM Tris-HCl و 500 mM KCl و 2 mM MgCl 2 و 2 mM CaCl 22 mM DTT و 5 mM ATP. - قم بتسمية الأنبوب على أنه APB وقم بتخزينه عند 4 درجات مئوية.
- امزج 455 ميكرولتر من المخزن المؤقت FC ، و 25 ميكرولتر من 100 mM ATP ، و 20 ميكرولتر من 50 mM DTT في أنبوب جهاز طرد مركزي دقيق.
- إعادة تشكيل الأكتين
- إعادة تشكيل الأكتين العضلي الهيكلي للأرانب عن طريق إضافة 100 ميكرولتر من الماء منزوع الأيونات إلى قارورة 1 ملغ من الأكتين المجمد بالتجميد. اخلطي جيدا عن طريق السحب بلطف لأعلى ولأسفل. Aliquot في عينات 5 ميكرولتر ، وتجميد المفاجئة ، وتخزين 10 ملغ / مل أكتين أليكوتس في -80 درجة مئوية.
- إعادة تشكيل الأكتين العضلي الهيكلي للأرنب الحيوي عن طريق إضافة 20 ميكرولتر من ماء التناضح العكسي. Aliquot في عينات 5 ميكرولتر ، وتجميد المفاجئة ، وتخزين 1 ملغ / مل من الأكتين البيوتينيل أليكوتس عند -80 درجة مئوية.
- بلمرة الأكتين غير المصنفة مع تثبيت الفولويدين الرودامين
- قم بإذابة قارورة واحدة من 10 ملغم / مل من الأكتين واحتفظ بها على الجليد.
- قم بإعداد مخزن GAB الطازج ، وأضف 100 ميكرولتر من GAB إلى الأكتين أليكوت ، واخلطه عن طريق السحب بلطف لأعلى ولأسفل. احتضان الحل على الجليد لمدة 1 ساعة.
- إعداد APB الطازجة أثناء الحضانة. بعد الحضانة ، قم ببلمرة الأكتين إلى خيوط عن طريق إضافة 11 ميكرولتر من APB إلى محلول الأكتين. اخلطي جيدا عن طريق السحب بلطف لأعلى ولأسفل. يوضع على الجليد لمدة 20 دقيقة.
- أضف 5 ميكرولتر من الفلويدين المسمى بالرودامين إلى محلول خيوط الأكتين المبلمر الطازج. اتركيه على الجليد في الظلام لمدة 1 ساعة.
- قم بتخزين قارورة الأكتين الرودامين الملفوفة بورق الألومنيوم في الظلام عند 4 درجات مئوية.
ملاحظة: يقترح استخدام هذه الخيوط لمدة أقصاها 1 أسبوع. يمكن تأكيد جودة التركيز البؤري التلقائي كل يوم من خلال تصوير سريع لخلية تدفق تحتوي على التركيز البؤري التلقائي فقط وعرض خيوط متسقة يوما بعد يوم.
- بلمرة الأكتين البيوتينيل مع تثبيت اليكسا فلور 488 الفيلويدين
- قم بإذابة قارورة واحدة من 10 ملغم / مل من الأكتين وقارورة واحدة من 1 ملغ / مل من الأكتين الحيوي والاحتفاظ بها على الجليد.
- اصنع مخزن مؤقت GAB جديدا.
- اجمع بين القارورتين (الخطوة 2.8.1) في نسبة أكتين 10: 1: البيوتينيلات. أضف 100 ميكرولتر من GAB إلى خليط الأكتين واخلطه جيدا عن طريق السحب بلطف لأعلى ولأسفل. احتضان على الجليد لمدة 1 ساعة.
- جعل APB جديدة أثناء الحضانة.
- بعد خطوة الحضانة ، قم ببلمرة الأكتين بإضافة 11 ميكرولتر من APB إلى محلول الأكتين. تخلط جيدا عن طريق سحب لأعلى ولأسفل بلطف. احتضان على الثلج لمدة 20 دقيقة.
- أضف 5 ميكرولتر من الفاليودين المسمى Alexa Fluor 488 واحتضنه على الثلج في الظلام لمدة 1 ساعة.
- قم بتخزين قارورة الأكتين البيوتينيل الملفوفة بورق الألومنيوم في الظلام عند 4 درجات مئوية.
ملاحظة: يمكن استخدام هذه الخيوط لمدة أقصاها 1 أسبوع.
3. إعداد الميوسين والخرز
- إعادة تشكيل ميوسين الثاني
- قم بتدوير لفترة وجيزة لأسفل (~ 5 ثانية) الميوسين الهيكلي المجفف بالتجميد II لجمعه في الجزء السفلي من الأنبوب باستخدام جهاز طرد مركزي صغير قياسي.
- أعد تشكيل الميوسين إلى 10 ملغم / مل عن طريق إضافة 100 ميكرولتر من 1 mM DTT المحضر في ماء RO.
- قم بتخفيف محلول الميوسين 10x عن طريق إضافة 10 ميكرولتر من 10 مجم / مل ميوسين إلى 90 ميكرولتر من 1 mM DTT في ماء RO. اصنع أليكوتات صغيرة الحجم (1-5 ميكرولتر) ، وقم بتجميدها ، وتخزينها عند -80 درجة مئوية.
ملاحظة: يمكن تأكيد نشاط الميوسين عن طريق إجراء فحص خيوط الانزلاق القياسي كما هو منشور سابقا46,47. انظر المناقشة للحصول على وصف موجز.
- تنظيف الخرز المطلي بالستربتافيدين
- تمييع 20 ميكرولتر من 1 ميكرومتر من حبات الستربتافيدين إلى 80 ميكرولتر من ماء التناضح العكسي. يغسل أربع مرات عن طريق الدوران لأسفل عند 9,600 × جم وإعادة تكوينه في 100 ميكرولتر من ماء التناضح العكسي.
- سونيك لمدة 2 دقيقة بسعة 40٪ وتخزين الخرز المغسول على دوار عند 4 درجات مئوية.
4. إعداد خلية التدفق
- تحضير محلول بولي ل ليسين (PLL) عن طريق إضافة 30 مل من الإيثانول 100٪ إلى أنبوب 50 مل وإضافة 200 ميكرولتر من 0.1٪ ث / v بولي ل ليسين في الماء وتخلط جيدا.
- أضف غطاء محفورا إلى محلول PLL واتركه ينقع لمدة 15 دقيقة. قم بإزالة الغطاء باستخدام ملاقط ، مع الحرص على لمس حافة الغطاء فقط أثناء سحبه من الأنبوب (انظر الشكل 1A-C). أمسك بأغطية الغطاء من حوافها بيد قفازات.
- جفف الغطاء مع شركة طيران مفلترة حتى لا يكون هناك إيثانول متبق ولا بقايا على الغطاء.
- ضع قطعتين من الشريط اللاصق على الوجهين في منتصف شريحة المجهر ، على بعد 3-4 مم من بعضها البعض. قم بتمزيق أو قطع الشريط الزائد الذي يتدلى من حافة الشريحة.
- أضف الغطاء المطلي ب PLL أعلى الشريط عموديا على المحور الطويل لشريحة المجهر (التي تشكل T) لتشكيل قناة.
- استخدم أنبوبا صغيرا لضغط الغطاء على الشريط وشريحة المجهر جيدا حتى يصبح الشريط شفافا (الشكل 1A). تأكد من عدم وجود فقاعات في الشريط لأن هذا يمكن أن يسبب تسربا من قناة التدفق.
ملاحظة: يمكن لخلية التدفق الاحتفاظ بحجم 10-15 ميكرولتر.
5. إعداد حزمة أكتوميوسين
- في أنابيب منفصلة ، قم بتخفيف كل نوع من خيوط الأكتين (الرودامين والبيوتينيل 488 الموسومة) 600x عن طريق خلط 0.5 ميكرولتر من الأكتين المحترم المسمى مع 300 ميكرولتر من APB. أضف 5 ميكرولتر إضافية من الفيلويدين المسمى بالمقابل إلى كل أنبوب واحتضنه على الثلج في الظلام لمدة 15 دقيقة.
- إلى محلول الأكتين البيوتينيل ، أضف نظام كسح الأكسجين من 1 ميكرولتر من بيتا دي الجلوكوز عند 500 ملغ / مل ، و 1 ميكرولتر من أوكسيديز الجلوكوز عند 25 ملغ / مل ، و 1 ميكرولتر من الكاتالاز عند 500 وحدة / مل. أضف 1 ميكرولتر من 100 mM ATP و 1 ميكرولتر من حبات الستربتافيدين المخففة والنظيفة 100x. يحرك بلطف بطرف ماصة. ضع التعليق على دوار عند 4 درجات مئوية بينما يتم تجميع بقية حزمة actomyosin.
- أضف 15 ميكرولتر من أكتين الرودامين المخفف إلى خلية تدفق PLL (الشكل 1D). قم بفتيل المحلول الزائد من خلال خلية التدفق ولكن لا تسمح لقناة التدفق بأن تصبح جافة. احتضان لمدة 10 دقائق في غرفة الرطوبة.
ملاحظة: يمكن صنع غرف الرطوبة من صناديق أطراف ماصة فارغة مع إضافة الماء إلى الأسفل والغطاء المغطى بورق الألومنيوم لمنع الضوء. - تحضير محلول الكازين 1 ملغ / مل في APB.
- يضاف 15 ميكرولتر من الكازين 1 ملغم/مل لمنع الربط غير المحدد للمكونات اللاحقة (الشكل 1 هاء). احتضان لمدة 5 دقائق في غرفة الرطوبة.
- أضف التركيز المطلوب من الميوسين إلى الأكتين الحيوي وتعليق الخرز من الخطوة 5.2. حرك بلطف بطرف الماصة ، ثم أضف على الفور 15 ميكرولتر من تعليق الخطوة 5.2 + تركيز الميوسين المطلوب إلى خلية التدفق (الشكل 1F ، G). احتضان لمدة 20 دقيقة. أغلق الأطراف المفتوحة لخلية التدفق باستخدام طلاء الأظافر لمنع التبخر أثناء التصوير وتجارب الاصطياد البصري.
ملاحظة: ينتج عن تركيز محلول الميوسين البالغ 1 ميكرومتر تجميعا قويا ويمكن استخدامه كنقطة انطلاق للتخصيص المطلوب للفحص (انظر الشكل 2).
6. قياسات القوة باستخدام المصيدة البصرية (NT2 Nanotracker2)
ملاحظة: على الرغم من أن البروتوكول أدناه مخصص لنظام NT2 ، إلا أنه يمكن استخدام هذا الفحص مع أنظمة الملائمة البصرية الأخرى ، بما في ذلك تلك المصممة خصيصا ، والتي لديها أيضا قدرات التألق. يظل سير العمل العام كما هو في التركيز البؤري على سطح الشريحة ، وإجراء معايرات الخرز ، والحصول على البيانات من خلال العثور على حزم الأكتين الفلورية. وبالنسبة لنظام NT2، يقدم الشكل التكميلي S1، والشكل التكميلي S2، والشكل التكميلي S3، والشكل التكميلي S4، والشكل التكميلي S5، والشكل التكميلي S6، والشكل التكميلي S7 تفاصيل عن نظام الاصطياد البصري وواجهة البرامجيات.
- قم بتشغيل مربع التحكم والليزر (الشكل التكميلي S1).
- ابدأ تشغيل برنامج كمبيوتر الملائمة البصرية بالنقر فوق رمز JPK Nanotracker على سطح المكتب.
- قم بإيقاظ وحدة التحكم عن بعد بالنقر فوق الزر Logitech في الوسط (الشكل التكميلي S2).
- قم بتشغيل وحدة التألق عن طريق تبديل مفتاح التشغيل/الإيقاف (الشكل التكميلي S3).
- أدر برج مكعب المرشح للتصوير الساطع (الشكل التكميلي S4).
- بمجرد أن يصبح النظام جاهزا ، قم بتشغيل الليزر باستخدام زر طاقة الليزر في الزاوية السفلية اليسرى من الشاشة إلى 50 ميجاوات واتركه يستقر لمدة 30 دقيقة (الشكل التكميلي S5).
- انقر بالتتابع على أزرار الإضاءة والكاميرا والهدف وحركة المسرح داخل البرنامج لإظهار تلك النوافذ للعرض والتلاعب أثناء التجربة. قم بتشغيل إضاءة المجهر من خلال النقر فوق الزر تشغيل / إيقاف تشغيله وضبطه على أقصى طاقة عن طريق النقر فوق الشريط وسحبه على طول الطريق إلى اليمين (الشكل التكميلي S5).
- افتح منطقة العينة وأزل حامل العينة من مرحلة المجهر. أضف خلية التدفق ، وقم بتأمينها باستخدام حاملات العينات المعدنية ، وتأكد من أن الشريحة مع الغطاء في الأسفل.
- أضف 30 ميكرولتر من ماء RO إلى وسط الهدف السفلي. لا تدع طرف الماصة يلمس العدسة. أعد إدراج مرحلة العينة.
ملاحظة: نظرا لأن نظام NT2 يستخدم هدف غمر الماء كهدف ملائمة، فقد تختلف وسائط الغمر وفقا لهدف الملائمة في إعداد المستخدم. - ارفع الهدف السفلي باستخدام أسهم التحكم على الشاشة أو L2 على وحدة التحكم عن بعد حتى تلامس حبة الماء الغطاء (الشكل التكميلي S5).
- اخفض الهدف العلوي حتى يتم الوصول إلى حوالي نصف المسافة إلى خلية التدفق باستخدام الأسهم التي تظهر على الشاشة أو R2 على وحدة التحكم عن بعد. أضف 170 ميكرولتر من ماء RO إلى أعلى خلية التدفق مباشرة تحت الهدف العلوي. اخفض الهدف العلوي حتى يكسر التوتر السطحي للماء ويشكل غضروفا مفصليا.
- حرك مرحلة المجهر باستخدام لوحة الأسهم الموجودة على وحدة التحكم عن بعد حتى يتم الوصول إلى حافة الشريط المجاور لقناة التدفق. أغلق باب العينة.
ملاحظة: تشير "النقرة" عند إغلاق باب العينة إلى أن غالق الليزر مفتوح الآن. هذه ميزة أمان تسمح فقط بفتح الغالق إذا كان الباب مغلقا. - باستخدام نافذة الهدف في الشاشة، ضع حافة الشريط في بؤرة التركيز البؤري عن طريق رفع الهدف السفلي المسمى هدف الليزر لأعلى من خلال النقر على السهم العلوي باستخدام عناصر التحكم التي تظهر على الشاشة. افعل الشيء نفسه بالنسبة للهدف العلوي بالنقر فوق السهم السفلي (الشكل التكميلي S5).
ملاحظة: تحرك الأسهم المزدوجة الهدف أو المرحلة بشكل أسرع. يتم استخدام حافة الشريط للتركيز البؤري لأنه كائن كبير يسهل العثور عليه وقريب من سطح الغطاء. فقاعات الهواء داخل الشريط هي خيار آخر. ومع ذلك ، هذا غير مطلوب إذا كان لدى المستخدم روتين تلقائي للعثور على التركيز السطحي أو طريقة داخلية مفضلة. - بمجرد أن يصبح الشريط في بؤرة التركيز، أغلق القزحية جزئيا في الجزء العلوي من المصيدة البصرية. اخفض الهدف العلوي حتى يظهر شكل مضلع القزحية. ضع هذه الحواف في بؤرة التركيز ، وأعد فتح القزحية ، ثم قم بإقران الأهداف باستخدام النقر فوق رمز القفل (الشكل التكميلي S5).
- ابحث عن حبة عائمة وقم باحتجازها بالنقر فوق زر Trap Shutter ، والذي سيفتح الغالق ويسمح لليزر الاصطياد بضرب العينة. انقر على مؤشر ملائمة على الشاشة واسحبه لنقل موقع ليزر الملائمة. بمجرد محاصرتها ، قم بمعايرة الخرزة لربط قياسات الجهد بالقوة والإزاحة.
- انقر على زر المعايرة . اضبط روتين المعايرة استنادا إلى تحليل أطياف الطاقة وقم بملاءمة تردد الزاوية داخل البرنامج لاتجاهات X و Y و Z (الشكل التكميلي S6).
- انقر على الإعدادات. اكتب قطر الخرزة (1000 نانومتر) ، واكتب درجة حرارة المرحلة الموجودة في أسفل يسار نافذة البرنامج. (انظر الشكل التكميلي S6).
- انقر على فخ 1. انقر على X Signal. انقر فوق تشغيل لأداء تناسب تردد الزاوية. انقر واسحب داخل النافذة لتحسين ملاءمة الوظيفة. انقر فوق استخدامه للحصول على قيم الحساسية والصلابة. انقر على قبول القيم. كرر ذلك لإشارات Y و Z. أغلق النافذة. (انظر الشكل التكميلي S6).
ملاحظة: إجراءات معايرة الخرز على أنظمة الملائمة البصرية الأخرى أو الأنظمة المصممة خصيصا والتي تم اختبارها بقوة من قبل المستخدم ، مثل طريقة التقسيم المتساوي أو طريقة قوة السحب ، مقبولة أيضا57,58. - ابحث عن حزمة أكتوميوزين من خلال البحث عن الخرز المرتبط بالتركيز البؤري التلقائي على سطح الغطاء.
- عندما يتم الكشف عن خرزة غير مزدحمة بالخرز العائم الآخر ، راقب التركيزات البؤرية التلقائية المحيطة بها عن طريق التصوير الفلوري للتحقق من وجود حزمة.
- تحقق من وجود حزمة من خلال البحث عن كل من AFs الفلورسنت المترجمة معا. قم بتشغيل مصدر الضوء الأبيض واستخدم مكعب المرشح المناسب لتصوير كل خيوط الأكتين عن طريق تدوير البرج (مكعبات مرشح الإثارة 488 نانومتر و 532 نانومتر لإثارة Alexa Fluor 488 و rhodamine ، على التوالي). انظر الشكل التكميلي S4.
ملاحظة: يمكن أن تكون تجربة التحكم للتحقق من شدة التألق للتركيز البؤري التلقائي الفردي مفيدة في تحديد الحزم التي تتكون من خيوط واحدة تحمل علامة 488 و 8 من الرودامين ، أو تنطبق على أي مجموعة من الفلوروفورات التي يختار المستخدم استخدامها. - بمجرد التحقق ، قم بحبس الخرزة المرفقة بالخيوط العلوية للحزمة بالنقر فوق الزر Trap Shutter .
- استخدم عناصر التحكم التي تظهر على الشاشة لتسجيل البيانات بالنقر فوق زر الذبذبات (الشكل التكميلي S7). لتصور القياسات دون تسجيل البيانات ، انقر فوق ابدأ. لحفظ جميع البيانات ، انقر فوق الحفظ التلقائي. لتسجيل القياسات، انقر فوق بدء السجل. اختر البيانات التي سيتم تصورها في الوقت الفعلي (الموضع ، القوة ، اتجاه x ، اتجاه y) عن طريق الاختيار من القائمة المنسدلة X إشارة أو إشارة Y. تذكر أن xdirection من اليسار إلى اليمين، والاتجاه y لأعلى ولأسفل على الشاشة. انظر الشكل التكميلي S7.
ملاحظة: سيتم حفظ البيانات كملفات .out وتتضمن الوقت والجهد والإزاحة والقوة لكل اتجاه. يمكن تصدير هذه الملفات إلى برامج أخرى للتصور والتحليل.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
تتميز خلايا التدفق التي تحتوي على أنظمة حزم الأكتوميوزين بتصميم قياسي ، وتتكون من شريحة مجهرية وغطاء محفور مفصول بقناة مصنوعة من شريط لاصق على الوجهين (الشكل 1). ثم يتم بناء الفحص من الغطاء إلى الأعلى باستخدام مقدمات مرحلية كما هو موضح في البروتوكول. يتكون الفحص النهائي من خيوط الأكتين الموسومة بالرودامين النموذجية. تركيز الميوسين المطلوب (تم استخدام 1 ميكرومتر للنتائج التمثيلية في الشكل 2 والشكل 3) ؛ البيوتينيل ، خيوط الأكتين اليكسا فلور 488 ؛ 1 ميكرومتر من حبات الستربتافيدين ؛ نظام كسح الأكسجين. رابطة محترفي التنس. والمخزن المؤقت APB. سيتم تشكيل حزم متعددة لكل خلية تدفق ، وتركيزات الأكتين الموضحة أعلاه تعطي تباعدا كافيا بين الحزم لضمان عدم وجود تفاعلات غير مرغوب فيها. وهذا يسهل أيضا الحصول على قياسات قوة متعددة لكل خلية تدفق لزيادة كفاءة الحصول على البيانات. يجب أن تكون ملفات تعريف القوة قابلة للتكرار داخل خلية التدفق ومن خلية التدفق إلى خلية التدفق.
في حين أن البروتوكول أعلاه موجه نحو استخدام إعداد الاصطياد البصري التجاري ، يمكن بسهولة استخدام خلية التدفق والفحص المعروضين هنا لأداة تجارية مختلفة أو إعداد محاصرة بصرية مصمم خصيصا إلى جانب مرحلة المجهر أو المجهر وامتلاك قدرات التصوير الفلورية. بمجرد اكتمال جميع إضافات خلايا التدفق وفقا للبروتوكول أعلاه ، تكون حزم actomyosin الموجودة على الشريحة (الشكل 1) جاهزة للقياس الفوري. تتم إضافة خلية التدفق إلى مرحلة مجهر المصيدة الضوئية ، ويتم الحصول على قياسات معايرة حبات متعددة ، ويتم تحديد الحزم من خلال التوطين المشترك الفلوري لخيوط الحزمة. يتم حبس حبسة مرتبطة بحزمة ، وتبدأ الإزاحة وقياس القوة المقابلة. يمكن للمستخدم مراقبة الحصول على البيانات في الوقت الحقيقي على شاشة الكمبيوتر. اعتمادا على تركيز الميوسين المستخدم في خلية التدفق ، يمكن أن تبدأ الحزمة في إظهار حركة كبيرة على الفور ، أو قد يستغرق الأمر 30 s-1 min لرؤية زيادة في الإزاحة / القوة بشكل فعال.
يظهر أثر القوة التمثيلية في الشكل 3A حيث تظهر محركات الميوسين منحدرا ثابتا في القوة متبوعا بهضبة. من المعتاد أن نرى هذه الأنواع من الآثار تتطور على مدى 2-5 دقائق. ومع ذلك ، من الممكن أيضا قياس حزم الأكتوميوزين التي لا تولد أي قوة صافية (الشكل 3B). تظهر هذه الآثار كضوضاء أساسية أو لا تظهر أي زيادة صافية كبيرة في القوة على مدى 90 ثانية. من المحتمل أن يكون هذا بسبب انخفاض التركيز المحلي للمحرك الذي لا يسمح بالانزلاق الإنتاجي ، أو أن الحزمة في اتجاه متوازي غير موات حيث يتم محاذاة نهايات الزائد والناقص للخيوط.
نظرا لأن محتويات خلية التدفق يمكن أن تكون عرضة للتدهور من الإضاءة الحادثة وليزر الاصطياد ، والتسخين المحلي على الشريحة بمرور الوقت ، وتوليد أنواع الأكسجين الجذرية ، ينصح بشدة بعدم استخدام نفس خلية التدفق لأكثر من 1 ساعة. لتحقيق أقصى قدر من الكفاءة ، يقترح إجراء فحص آخر أثناء الحصول على البيانات. يمكن تصدير تتبع الإزاحة / القوة من برنامج الملائمة البصرية إلى Excel أو Matlab أو Igor أو برامج إدارة البيانات الأخرى لمزيد من التصفية والتحليل. تتضمن البيانات التي يمكن استخراجها من تجارب مجموعة/حزمة الاصطياد البصري هذه أنواعا مختلفة من ملفات تعريف توليد القوة (خط الأساس، المنحدر/الهضبة) في ظل ظروف مقايسة مختلفة، وسرعة توليد القوة، والحد الأقصى لتوليد القوة، والسلوك الحركي وسلوك المجموعة من خلال أحجام الخطوات وأوقات السكن بين الخطوات أو فرق الخطوات، فضلا عن نسبة الواجب. يمكن للمستخدم أيضا تغيير شروط الفحص لمقارنة كيفية تأثير إضافة أنواع مختلفة من محركات الميوسين أو إضافة بروتينات ربط الأكتين أو تغيير ظروف التخزين المؤقت على خصائص توليد قوة المجموعة هذه.
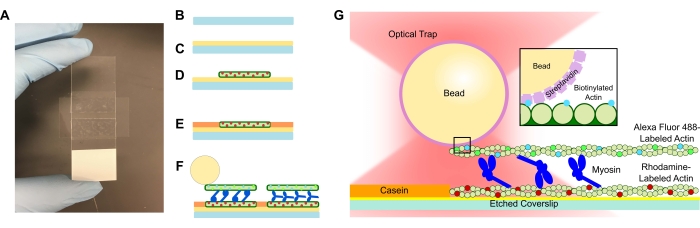
الشكل 1: الفحص التخطيطي (A-C) يتم طلاء الأغطية المحفورة بالبولي L-lysine وتستخدم لتشكيل خلية التدفق باستخدام شريط على الوجهين وشريحة مجهرية. تؤدي المقدمات الموقوتة وخطوات الحضانة الموصوفة في البروتوكول إلى الأكتين المستقر بالفلويدين المسمى بالرودامين كقالب أو خيوط سفلية (D) ، يليه حجب الكازين لمنع الربط غير المحدد (E) ، و (F) Alexa Fluor 488 الأكتين البيوتينيل المستقر بالفيلويدين كشحنة أو خيوط علوية ، وفرق من الميوسين II التي تفصل الخيوط عن بعضها البعض وتولد القوة عند إدخال ATP. يمكن أن تختلف هندسة المحركات وطبيعة الربط المتقاطع داخل الحزمة في ظل ظروف مختلفة ، مثل تركيز الملح59. وقد أظهرت الدراسات السابقة أن مجال ذيل الميوسين لديه القدرة على التفاعل مع خيوط الأكتين وحركية المجموعة البطيئة46. ومع ذلك ، فإن رؤوس الميوسين في تجارب الميروميوسين الثقيلة تظهر ارتباط كل رأس بخيوط الأكتين المجاورة60. (ز) تستخدم حبات الستربتافيدين كمقبض بصري للمصيدة وترتبط فقط بخيوط الأكتين البيوتينيل البضائع، مما يساعد في التحقق من تكوين الحزم المناسبة على الشريحة. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.

الشكل 2: حزم أكتوميوسين الفلورسنت. أربعة لقاءات مختلفة لخيوط الأكتين والحزم داخل فحص الحزمة المعروضة في الشكل 1. يتم عرض خيوط الأكتين البيوتينيل العلوية للبضائع مع قناة Alexa Fluor 488 phalloidin على اليسار ، وخيوط الأكتين السفلية مع قناة phalloidin rhodamine على اليمين. في الجزء السفلي ، يظهر نفس الشكل مع خطوط ملونة متراكبة للمساعدة في توجيه العين. (أ) تم العثور على خيوط أكتين علوية بالقرب من خيوط الأكتين السفلية ولكن لها تراكب غير مكتمل. لن يتم استخدام هذا لتجارب الحزمة. (ب) يتم تجميع خيوط الأكتين العلوية والسفلية بشكل مشترك، وتؤكد شدة كل خيط أنها خيوط واحدة داخل الحزمة. سيكون هذا مرشحا جيدا لتجارب الحزمة. (ج) توجد حزمة كبيرة من خيوط الرودامين ذاتية التجميع في القاع. في حين أن هناك خيوط أكتين علوية مقابلة يتم توطينها بشكل مشترك ، إلا أن هناك الكثير من الخيوط السفلية الموجودة. وبالتالي ، لن يتم استخدامه لتجارب الحزمة. هذا أيضا مثال على كيفية زيادة كثافة التألق عندما يتم تجميع خيوط أكتين متعددة من نفس النوع. يمكن للمستخدم استخدام هذا كمقياس للحكم على الخيوط الفردية مقابل الحزم من نفس نوع الخيوط. (د) توجد خيوط سفلية بدون خيوط علوية مقابلة، مما يؤكد أيضا عدم وجود نزف. لن يتم استخدام هذا لتجارب الحزمة. نلاحظ أن كثافة الخيوط في قناة Alexa Fluor 488 منخفضة ونعتقد أنها ترجع إلى مجموعة المرشحات المستخدمة (Filter Set 09 من Zeiss). مجموعة المرشحات المستخدمة لقناة الرودامين هي مجموعة التصفية 43 من زايس. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.

الشكل 3: توليد قوة مجموعة Myosin II. آثار تمثيلية لمحركات الميوسين II الهيكلية التي تولد القوة داخل التسلسل الهرمي الهيكلي للأكتين الذي تم بناؤه في المختبر . تعمل محركات الميوسين معا لتوليد القوة بشكل جماعي ومنتج حتى يتم الوصول إلى هضبة واستمرار القوة (A) أو تجربة العداء بالقرب من خط الأساس (B). يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
الشكل التكميلي S1: مصيدة بصرية Bruker / JPK Nanotracker2. (أ) شاشة الكمبيوتر. (ب) لوحة مفاتيح الكمبيوتر. (ج) برج الحاسوب. (د) مربع تحكم. (ه) إمدادات الطاقة بالليزر. (و) صندوق بصريات الفخ البصري. (ز) المجهر المقلوب. (ح) مرحلة الباب إلى المجهر. (I) شريط تمرير المستقطب للتبديل بين التصوير الساطع للتداخل والتفاضلي. يرجى النقر هنا لتنزيل هذا الملف.
الشكل التكميلي S2: جهاز تحكم عن بعد للمصيدة البصرية. (أ) لوحة مفاتيح لوضع المرحلة الآلية. (ب-ج) اضبط موضع الملائمة. (D) تشغيل A و X و B وإيقاف تشغيل الغالق الرئيسي ، ومصراع فخ 1 ، ومصراع فخ 2 ، على التوالي. (ه) يستخدم زر Logitech لإيقاظ وحدة التحكم. (و) الزران لأعلى ولأسفل المستخدمان لوضع هدف الملائمة. (ز) الزران لأعلى ولأسفل المستخدمان لوضع هدف الكشف. لاحظ أن جهاز التحكم عن بعد غير مطلوب ، ويمكن إنجاز كل هذه التلاعب في البرنامج. ومع ذلك ، فمن الملائم أن تكون قادرا على التحكم في الأهداف وموضع المرحلة أثناء النظر في بيئة مرحلة المجهر. يرجى النقر هنا لتنزيل هذا الملف.
الشكل التكميلي S3: وحدة التألق للمصيدة البصرية. يقترن مصدر الضوء الأبيض الفلوري 89North PhotoFluor بالجزء الخلفي من المجهر المقلوب. يتم تشغيله وإيقاف تشغيله باستخدام مفتاح تبديل (سهم). يرجى النقر هنا لتنزيل هذا الملف.
الشكل التكميلي S4: برج مكعب مرشح التألق. يمكن تشغيل البرج (السهم) لاستخدام مكعب المرشح الضروري للتصوير في أصباغ DIC أو الرودامين أو Alexa Fluor 488. لاحظ أنه يمكن تبديل مكعبات المرشح لتخصيص الإعداد لاستخدام الفلوروفورات المختلفة. يرجى النقر هنا لتنزيل هذا الملف.
الشكل التكميلي S5: برنامج Nanotracker2. (أ) زر الطاقة بالليزر والتحكم. (ب) نافذة تحديد المواقع الموضوعية. تستخدم الأسهم الاتجاهية لنقل أهداف الكشف (أعلى) والملائمة (أسفل). الأسهم المزدوجة تحرك الأهداف بسرعة أعلى. الزر الأزرق والأحمر في أسفل اليسار يفصل بين الأهداف ويسحبها مرة أخرى إلى موضعها الأصلي. هذا ضروري عند أخذ عينات داخل وخارج مرحلة المجهر. الزر الثالث من اليسار مع الأهداف ورمز القفل "يزوج" الأهداف بحيث عندما تكون في بؤرة التركيز وتحقيق إضاءة كوهلر ، يمكن للمستخدم تحريك كل من أهداف الاصطياد والكشف لأعلى ولأسفل في المحور z. (ج) نافذة تحديد المواقع للعينة المستخدمة لتحريك مرحلة المجهر في المحورين x و y. الأسهم المزدوجة تحرك المرحلة بسرعة أعلى. يتم تنشيط هذه النافذة بالنقر فوق رمز السهم لأعلى / لأسفل ولليسار / لليمين في القائمة العلوية. (د) نافذة تصور الكاميرا. يمكن استخدام رمز مفتاح الربط لتعيين ظروف التصوير المخصصة. يتم تنشيط هذه النافذة بالنقر فوق رمز الكاميرا في القائمة العلوية. (ه) نافذة إضاءة المجهر. يتم تنشيط هذه النافذة بالنقر فوق رمز المصباح الكهربائي في القائمة العلوية. يرجى النقر هنا لتنزيل هذا الملف.
الشكل التكميلي S6: نافذة المعايرة. (أ) تستخدم هذه النافذة لمعايرة الخرز ويتم تنشيطها بالنقر فوق أيقونة Cal في القائمة العلوية. لمعايرة حبة ، يتم تحقيق أفضل ملاءمة لتردد الزاوية في إشارات x و y و z. (ب) لكل إشارة، اختر زر الإشارة المناسب في أعلى اليسار. (C) انقر فوق تشغيل وتحسين الملاءمة عن طريق النقر والسحب داخل النافذة الخضراء (D). (ه) بمجرد الرضا عن الملاءمة ، انقر فوق استخدامه للحساسية والصلابة. سيسمح ذلك بتسجيل الإزاحة بالنانومتر والقوة في البيكونوتون. (F) ثم انقر فوق قبول القيم في أسفل اليسار. كرر ذلك للاتجاهين y و z. يرجى النقر هنا لتنزيل هذا الملف.
الشكل التكميلي S7: نافذة الحصول على البيانات. تستخدم هذه النافذة للحصول على بيانات الموقع والقوة وتسمح للمستخدم برؤية القياسات في الوقت الفعلي. (أ) يتم تنشيط هذه النافذة بالنقر فوق الرمز x,t في القائمة العلوية. (B) يمكن للمستخدم التبديل بين عرض إشارات x و y. (ج) انقر فوق ابدأ لبدء تصور البيانات. انقر على الحفظ التلقائي لحفظ البيانات. انقر فوق بدء التسجيل لبدء تسجيل البيانات وحفظها. يرجى النقر هنا لتنزيل هذا الملف.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
تم إجراء دراسة في المختبر باستخدام ملاقط بصرية جنبا إلى جنب مع التصوير الفلوري للتحقيق في ديناميكيات مجموعات الميوسين التي تتفاعل مع خيوط الأكتين. تم تجميع حزم الأكتين-الميوسين-الأكتين باستخدام الميوسين العضلي الثاني، والأكتين الرودامين في الجزء السفلي من الحزمة وعلى سطح الغطاء، وخيوط الأكتين البيوتينيل ذات العلامات 488 في الجزء العلوي من الحزمة. تم بلمرة بروتين الأكتين من عضلة الأرانب وتثبيته باستخدام مخازن الأكتين العامة (GAB) ومخازن بلمرة الأكتين (APB). يجب إعداد GAB و APB طازجا كل يوم في المختبر باستخدام ATP و FC buffer و TC buffer. تم استخدام عضلات الميوسين II لتشكيل سندويشات الأكتين-ميوسين-أكتين. تم استخدام Phalloidin لتلطيخ الفلورسنت من خيوط الأكتين ، وكذلك الاستقرار في المختبر.
يمكن تأكيد نشاط الميوسين من خلال إجراء فحص خيوط الانزلاق القياسي كما هو منشور سابقا46,47. يمكن أن يرتبط الميوسين II وشظاياه الفرعية بسطح الغطاء في مجموعة متنوعة من الاتجاهات ، ويمكن أن يؤدي وجود مجال الذيل إلى إبطاء انزلاق الخيوط مقارنة بالفحوصات باستخدام الميروميوسين الثقيل 46،48،49. ومع ذلك ، لا يزال من الممكن ملاحظة الانزلاق والحركة السطحية. هناك دليل أكثر وضوحا على نشاط الميوسين هو كسر خيوط الأكتين النشط الذي يمكن ملاحظته حيث يتم تقسيم خيوط الأكتين الأطول إلى شظايا أصغر ثم تنزلق بعيدا في اتجاهات متعددة. يحدث هذا بسبب التركيز العالي للمحركات النشطة على السطح ، وقد لوحظت من قبل مختبرات متعددة ، ولا يحدث بدون محركات الميوسين النشطة الموجودة42،50،51،52،53،54. علاوة على ذلك ، يساعد فحص الحزمة المقدم هنا في التخفيف من مشكلات الحركة التي ارتبطت في المقام الأول بفحص الخيوط المنزلقة ، مثل مجموعة متنوعة من اتجاهات ربط المحرك على غطاء زجاجي ، لأن فحص الحزمة يتضمن حجب الكازين لسطح الزجاج بحيث ترتبط المحركات داخل الحزمة 47,55,56.
الخطوة الأولى هي إضافة خيوط الأكتين الرودامين كخيوط سفلية أو قالب إلى غطاء مغلف بالبولي إل-ليسين في خلية تدفق. يستخدم Poly-L-lysine لتعزيز ربط الأكتين لأن poly-lysine مشحون إيجابيا بينما يحتوي الأكتين على شحنات سالبة وقد تم استخدامه في مستحضرات الفحص الخلوي السابق في المختبر 61,62,63. قبل تكوين الحزمة ، تمت إضافة تخفيفات مختلفة للأكتين إلى خلية تدفق لتحسين تركيز الأكتين. في هذه الحالة ، كان 600x من المخزون هو التخفيف الأمثل الذي أسفر عن عدد كاف من خيوط القالب لتشكيل الحزم ولكن مع تباعد كاف بحيث تم تخصيص الحزم. تم تنفيذ التخفيف باستخدام المخزن المؤقت APB. وأعقب إضافة الأكتين الرودامين طبقة من الكازين لحجب السطح وتجنب الربط غير المحدد. تم احتضان خلية التدفق لمدة 30 دقيقة وغسلها بعد الحضانة باستخدام المخزن المؤقت لغسل أي خيوط أكتين غير مقيدة. وأخيرا ، تمت إضافة مزيج من الميوسين ، والأكتين 488 / البيوتين ، والخرز المطلي بالستربتافيدين إلى خلية التدفق لتسهيل تكوين حزمة الأكتين الميوسين. يجب أن يكون تركيز الخرز بحيث يكون هناك ما يكفي لربط الحزم المرتبطة بالسطح ويكفي في التعليق لتسهيل المعايرة. ومع ذلك ، يمكن أن يسبب التركيز العالي جدا للخرز صعوبة أثناء تجارب الاصطياد بسبب سقوط الخرز المجاور في مصيدة الليزر وتعطيل القياس. تضاف محركات الميوسين إلى المجموعة مباشرة قبل حقنها في الشريحة بحيث لا تتجمع محركات الميوسين بشكل استباقي مع الحمولة أو خيوط الأكتين البيوتينيل العلوية وبالتالي تربط الرودامين السفلي بتجميع خيوط الأكتين البيوتينيلية.
نظام الاصطياد البصري NT2 هو فخ بصري تجاري مع مزيج من التباين الساطع والتداخل التفاضلي (DIC) وطرق التصوير فوق الفلورية. وهو مقترن بمجهر مقلوب من Zeiss AxioObserver 3 مع أهداف محاصرة واكتشاف الغمر المائي 100x / NA 1.46 و 63x / NA 1.0. وقد تم تجهيز النظام بقدرة ملائمة النقر والسحب لملائمة ليزر واحدة ويمكن استخدامها أثناء التصوير في أي من الطرائق المدرجة سابقا. يتم الكشف عن الحزم المشكلة وتأكيدها باستخدام التصوير الفلوري. وجود مصدر ضوء أبيض مع مكعبات مرشح مناسبة (GFP / FITC و TRITC / CY3) يسمح بالتبديل السريع بين تصوير الخيوط. تم التحقق من التأثيرات البؤرية التلقائية المشتركة من خلال تصور التأثيرات البؤرية التلقائية عند الأطوال الموجية المختلفة للإثارة قبل أخذ كل قياس للقوة باستخدام ملاقط بصرية. نظرا لأن الخيوط يمكنها التبييض الضوئي بسرعة حتى مع كاشف كسح الأكسجين ، يقترح أن يقوم الباحثون بتحسين معلمات التصور مثل الكثافة ووقت التعرض قبل إجراء تجارب الحزمة.
تم استخدام الاصطياد البصري لأخذ قياسات القوة ، باستخدام حبات الستربتافيدين في وجود ATP لربط خيوط أكتين البضائع البيوتينيل وتنشيط توليد قوة الميوسين كمحول قوة. تم استخراج بيانات الإزاحة والقوة مقابل الوقت التي تم الحصول عليها عن طريق الاصطياد البصري من برنامج الاصطياد لتحليلها. ومع ذلك ، يوفر برنامج الملائمة التجاري أيضا إجراءات تحليل يمكن استخدامها ، أو يمكن برمجة خوارزميات مخصصة في برامج أخرى من قبل المستخدم لتصور وتحليل بيانات الملائمة. في أنظمة الملائمة البصرية المخصصة ، قد يكون لدى المستخدم ليزر إثارة بدلا من مصدر ضوء أبيض مع مرشحات ، والتي هي أيضا مقبولة للاستخدام. علاوة على ذلك ، يمكن تغيير أصباغ الفلورسنت لتكون مناسبة للمعدات الحالية التي قد يمتلكها المستخدم إذا لم تتداخل أطياف الانبعاثات وتسبب نزيفا.
نلاحظ أن الفحص المقدم هو اختبار أساسي يمكن تخصيصه من قبل المستخدم اعتمادا على سؤال البحث الخاص به في مجال ميكانيكا مجموعة actomyosin. يمكن أيضا تطبيق سير العمل العام على أنظمة المجموعات الخلوية الهيكلية الأخرى في المختبر التي قد تكون ذات أهمية ، مثل اختبارات حزمة الأنابيب الدقيقة التي تشكل نماذج دنيا من المغزل الانقسامي 32,61,63,64,65,66. يمكن أن تشمل التعديلات على سبيل المثال لا الحصر تغيير تسميات الفلوروفور المناسبة للإعداد الحالي للمستخدم ؛ تغيير تركيز الميوسين أو البناء أو النمط المتماثل ؛ ومعايرة ظروف المخزن المؤقت، من بين جوانب أخرى.
التحديات المحتملة ممكنة عند إجراء هذا الفحص. عند تشكيل حزم الأكتين-الميوسين، قد لا يكون تركيز الميوسين داخل حزم الأكتين متجانسا عبر الشريحة. ولاستيعاب ذلك، سيتم قياس حزم متعددة عبر الشريحة بأكملها لضمان أخذ عينات من توزيع المحركات وتوليد القوة بشكل صحيح. ومن الصعب أيضا معرفة اتجاه الحزمة إذا كان ذلك مطلوبا لتفسير بيانات القوة. وبالتالي ، ينبغي إجراء تجارب متعددة لكل حزمة. يمكن للمرء أيضا دمج وضع العلامات النهائية لخيوط الأكتين من خلال الجيلسولين الفلوري أو الخرز المطلي بالجيلسولين بحجم أصغر من مقبض الاصطياد البصري. يمكن أيضا استخدام التصوير الفلوري للنظر في قوى المكون x و y لاستنتاج اتجاه الحزمة. علاوة على ذلك ، نظرا لأن حالة تراكم الميوسين تتأثر بشدة بالقوة الأيونية للمخزن المؤقت مع تكوين خيوط سميكة تحدث عند التخفيف السريع ل KCl ، يجب مراقبة تركيز الملح العازل بشكل مناسب67,68.
كانت الدراسات السابقة التي استخدمت طرقا أخرى في المختبر مثل مقايسات الانزلاق مفيدة لتحديد دور مجالات الميوسين ودراسة التكوين والتفاعلات بين الميوسين والبروتينات الأخرى المرتبطة بالأكتين. ومع ذلك ، فإن هذه الطرق لها عيب في أن ربط الميوسين على سطح صلب سيحد من إمكانية التنسيق بين محركات الميوسين وبالتالي ردود الفعل الميكانيكية التي تحدث لتحديد ما إذا كانت مجموعة المحرك في وضع نسبة واجب عالية أو منخفضة 33,35,41,69. علاوة على ذلك ، فإن الاصطياد البصري مع شبكات محركات الميوسين المفردة لا يعطي فهما واضحا لكيفية تفاعل محركات الميوسين مع بعضها البعض ومع خيوط الأكتين. يسمح البروتوكول الذي تم تطويره هنا بالتحقيق في ديناميكيات مجموعة محركات الميوسين داخل شبكة أكتين هرمية متوافقة. كما أنه قابل للتخصيص من حيث خصائص مجموعة خيوط المحرك مثل التركيز ، والشكل المتساوي ، والبيئة العازلة ، من بين جوانب أخرى ، للسماح بإجراء تحقيق منهجي. البروتوكول المقدم هو منصة للدراسات المستقبلية لشبكات الأكتوميوزين الأكثر تعقيدا ويحافظ على دقة قياسات الإزاحة وتوليد القوة التي يسهلها الاصطياد البصري الذي كان يستخدم تقليديا لدراسات الجزيء الواحد.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
ليس لدى المؤلفين أي تضارب في المصالح للإعلان.
Acknowledgments
يتم دعم هذا العمل جزئيا من قبل زمالة أبحاث مجلس طلاب الدراسات العليا بجامعة ميسيسيبي (OA) ، وجامعة ميسيسيبي سالي ماكدونيل باركسديل مع مرتبة الشرف (JCW ، JER) ، واتحاد منح الفضاء في ميسيسيبي تحت رقم المنحة NNX15AH78H (JCW ، DNR) ، وجمعية القلب الأمريكية تحت رقم المنحة 848586 (DNR).
Materials
| Name | Company | Catalog Number | Comments |
| Actin protein (biotin): skeletal muscle | Cytoskeleton | AB07-A | Biotinylated actin protein |
| Actin protein, rabbit skeletal muscle | Cytoskeleton | AKL99-A | Actin protein |
| Alexa Fluor 488 Phalloidin | Invitrogen | A12379 | Actin stabilizer and Alexa Fluor 488 stain |
| ATP | Fisher scientific | BP413-25 | Required for actin assembly and myosin motility |
| Beta-D-glucose | Fisher scientific | MP218069110 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Blotting Grade Blocker (casein) | Biorad | 1706404 | Used to block surface from non-specific binding |
| CaCl2 | Fisher scientific | C79500 | Calcium chloride, provides the necessary control over the dynamics of actin myosin network |
| Catalase | Fisher scientific | ICN10040280 | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| Coverslips | Fisher scientific | 12544C | Used to make flow cells |
| DTT | Fisher scientific | AC327190010 | Used for buffer preparation |
| Ethanol | Fisher scientific | A4094 | Regent used for cleaning coverslips |
| Glucose oxidase | Fisher scientific | 34-538-610KU | Part of oxygen scavenging system used to reduce photobleaching during fluorescence imaging |
| KCl | Fisher scientific | P217-500 | Used for buffer preparation |
| KOH | Fisher scientific | P250-1 | Used to etch coverslips and adjust buffer pH |
| MgCl2 | Fisher scientific | M33-500 | Used for buffer preparation |
| Microscope slides | Fisher scientific | 12-544-2 | Used to make flow cells |
| Myosin II protein: rabbit skeletal muscle | Cytoskeleton | MY02 | Full length myosin motor protein isolated from rabbit skeletal muscle |
| Nanotracker2 | Bruker/JPK | NT2 | Optical trapping instrument |
| Poly-l-lysine | Sigma-Aldrich | P8920 | Facilities adhesion of actin filaments onto glass surface of the coverslip |
| Rhodamine Phalloidin | Cytoskeleton | PHDR1 | Actin stabilizer and rhodamine fluorescent stain |
| Streptavidin beads, 1 μm | Spherotech | SVP-10-5 | Optical trapping handle |
| Tris-HCl | Fisher scientific | PR H5121 | Used for buffer preparation |
References
- Goldstein, L. S. Kinesin molecular motors: transport pathways, receptors, and human disease. Proceedings of the National Academy of Sciences of the United States of America. 98 (13), 6999-7003 (2001).
- Lee Sweeney, H., Holzbaur, E. L. F.
- O'Connell, C. B., Tyska, M. J., Mooseker, M. S. Myosin at work: Motor adaptations for a variety of cellular functions. Biochimica et Biophysica Acta - Molecular Cell Research. 1773 (5), 615-630 (2007).
- Kaya, M., Tani, Y., Washio, T., Hisada, T., Higuchi, H. Coordinated force generation of skeletal myosins in myofilaments through motor coupling. Nature Communications. 8, 1-13 (2017).
- Akhshi, T. K., Wernike, D., Piekny, A. Microtubules and actin crosstalk in cell migration and division. Cytoskeleton. 71 (1), 1-23 (2014).
- Brawley, C. M., Rock, R. S. Unconventional myosin traffic in cells reveals a selective actin cytoskeleton. Proceedings of the National Academy of Sciences of the United States of America. 106 (24), 9685-9690 (2009).
- Hartman, M. A., Spudich, J. A. The myosin superfamily at a glance. Journal of Cell Science. 125 (7), 1627-1632 (2012).
- Spudich, J. A., et al.
- Sommese, R. F., et al. Molecular consequences of the R453C hypertrophic cardiomyopathy mutation on human β-cardiac myosin motor function. Proceedings of the National Academy of Sciences of the United States of America. 110 (31), 12607-12612 (2013).
- Nag, S., et al. The myosin mesa and the basis of hypercontractility caused by hypertrophic cardiomyopathy mutations. Nature Structural & Molecular Biology. 24 (6), 525-533 (2017).
- Kawana, M., Sarkar, S. S., Sutton, S., Ruppel, K. M., Spudich, J. A. Biophysical properties of human b-cardiac myosin with converter mutations that cause hypertrophic cardiomyopathy. Science Advances. 3 (2), 1-11 (2017).
- Girolami, F., et al. Novel α-actinin 2 variant associated with familial hypertrophic cardiomyopathy and juvenile atrial arrhythmias: A massively parallel sequencing study. Circulation: Cardiovascular Genetics. 7 (6), 741-750 (2014).
- Debold, E. P., et al. Hypertrophic and dilated cardiomyopathy mutations differentially affect the molecular force generation of mouse α-cardiac myosin in the laser trap assay. American Journal of Physiology - Heart and Circulatory Physiology. 293 (1), 284-291 (2007).
- Barron, J. T.
- Duke, T. A. J.
- Vilfan, A., Duke, T.
- Huxley, A. F. Muscle structure and theories of contraction. Progress in Biophysics and Biophysical Chemistry. 7, 255-318 (1957).
- Huxley, H. E. Fifty years of muscle and the sliding filament hypothesis. European Journal of Biochemistry. 271 (8), 1403-1415 (2004).
- Kad, N. M., Kim, S., Warshaw, D. M., VanBuren, P., Baker, J. E. Single-myosin crossbridge interactions with actin filaments regulated by troponin-tropomyosin. Proceedings of the National Academy of Sciences of the United States of America. 102 (47), 16990-16995 (2005).
- Veigel, C., Molloy, J. E., Schmitz, S., Kendrick-Jones, J. Load-dependent kinetics of force production by smooth muscle myosin measured with optical tweezers. Nature Cell Biology. 5 (11), 980-986 (2003).
- Spudich, J. A.
- Simmons, R. M., Finer, J. T., Chu, S., Spudich, J. A. Quantitative measurements of force and displacement using an optical trap. Biophysical Journal. 70 (4), 1813-1822 (1996).
- Finer, J. T., Simmons, R. M., Spudich, J. Single myosin molecule mechanics: piconewton forces and nanometre steps. Nature. 368 (6467), 113-119 (1994).
- Kron, S. J., Uyeda, T. Q. P., Warrick, H. M., Spudich, J. A. An approach to reconstituting motility of single myosin molecules. Journal of Cell Science. 98, 129-133 (1991).
- Molloy, J. E., Burns, J. E., Kendrick-Jones, B., Tregear, R. T., White, D. C. S. Movement and force produced by a single myosin head. Nature. 378 (6553), 209-212 (1995).
- Ruegg, C., et al. Molecular motors: Force and movement generated by single Myosin II molecules. Physiology. 17 (5), 213-218 (2002).
- Nayak, A., et al. Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function. Journal of Biological Chemistry. 295 (20), 7046-7059 (2020).
- Dupuis, D. E., Guilford, W. H., Wu, J., Warshaw, D. M.
- Tyska, M. J., et al. Two heads of myosin are better than one for generating force and motion. Proceedings of the National Academy of Sciences of the United States of America. 96 (8), 4402-4407 (1999).
- Tyska, M. J., Warshaw, D. M.
- Finer, J. T., et al.
- Al Azzam, O., Trussell, C. L., Reinemann, D. N. Measuring force generation within reconstituted microtubule bundle assemblies using optical tweezers. Cytoskeleton. 78 (3), 111-125 (2021).
- Wagoner, J. A., Dill, K. A. Evolution of mechanical cooperativity among myosin II motors. Proceedings of the National Academy of Sciences of the United States of America. 118 (20), 2101871118 (2021).
- Walcott, S., Warshaw, D. M., Debold, E. P. Mechanical coupling between myosin molecules causes differences between ensemble and single-molecule measurements. Biophysical Journal. 103 (3), 501-510 (2012).
- Stewart, T. J., Murthy, V., Dugan, S. P., Baker, J. E. Velocity of myosin-based actin sliding depends on attachment and detachment kinetics and reaches a maximum when myosin-binding sites on actin saturate. Journal of Biological Chemistry. 297 (5), 101178 (2021).
- Hilbert, L., Cumarasamy, S., Zitouni, N. B., Mackey, M. C., Lauzon, A. M. The kinetics of mechanically coupled myosins exhibit group size-dependent regimes. Biophysical Journal. 105 (6), 1466-1474 (2013).
- Debold, E. P., Walcott, S., Woodward, M., Turner, M. A. Direct observation of phosphate inhibiting the Force-generating capacity of a miniensemble of myosin molecules. Biophysical Journal. 105 (10), 2374-2384 (2013).
- Kaya, M., Higuchi, H. Nonlinear elasticity and an 8-nm working stroke of single myosin molecules in myofilaments. Science. 329 (5992), 686-689 (2010).
- Pertici, I., et al. A myosin II nanomachine mimicking the striated muscle. Nature Communications. 9 (1), 1-10 (2018).
- Cheng, Y. S., De Souza Leite, F., Rassier, D. E. The load dependence and the force-velocity relation in intact myosin filaments from skeletal and smooth muscles. American Journal of Physiology - Cell Physiology. 318 (1), 103-110 (2020).
- Stam, S., Alberts, J., Gardel, M. L., Munro, E. Isoforms confer characteristic force generation and mechanosensation by myosin II filaments. Biophysical Journal. 108 (8), 1997-2006 (2015).
- Rastogi, K., Puliyakodan, M. S., Pandey, V., Nath, S., Elangovan, R. Maximum limit to the number of myosin II motors participating in processive sliding of actin. Scientific Reports. 6, 1-11 (2016).
- Debold, E. P., Patlak, J. B., Warshaw, D. M. Slip sliding away: Load-dependence of velocity generated by skeletal muscle myosin molecules in the laser trap. Biophysical Journal. 89 (5), 34-36 (2005).
- Albert, P. J., Erdmann, T., Schwarz, U. S. Stochastic dynamics and mechanosensitivity of myosin II minifilaments. New Journal of Physics. 16, (2014).
- Erdmann, T., Schwarz, U. S. Stochastic force generation by small ensembles of myosin II motors. Physical Review Letters. 108 (18), 1-5 (2012).
- Guo, B., Guilford, W. H. The tail of myosin reduces actin filament velocity in the in vitro motility assay. Cell Motility and the Cytoskeleton. 59 (4), 264-272 (2004).
- Miller-Jaster, K. N., Petrie Aronin, C. E., Guilford, W. H. A quantitative comparison of blocking agents in the in vitro motility assay. Cellular and Molecular Bioengineering. 5 (1), 44-51 (2012).
- Mansoon, A., Balaz, M., Albet-Torres, N., Rosengren, K. J. In vitro assays of molecular motors -- impact of motor-surface interactions. Frontiers in Bioscience. 13, 5732-5754 (2008).
- Persson, M., et al. Heavy meromyosin molecules extending more than 50 nm above adsorbing electronegative surfaces. Langmuir. 26 (12), 9927-9936 (2010).
- Kron, S. J., Spudich, J. A. Fluorescent actin filaments move on myosin fixed to a glass surface. Proceedings of the National Academy of Sciences of the United States of America. 83 (17), 6272-6276 (1986).
- Yanagida, T., Nakase, M., Nishiyama, K., Oosawa, F. Direct observation of motion of single F-actin filaments in the presence of myosin. Nature. 307 (5946), 58-60 (1984).
- Tsuda, Y., Yasutake, H., Ishijima, A., Yanagida, T. Torsional rigidity of single actin filaments and actin-actin bond breaking force under torsion measured directly by in vitro micromanipulation. Proceedings of the National Academy of Sciences of the United States of America. 93 (23), 12937-12942 (1996).
- Stewart, T. J., et al. Actin sliding velocities are influenced by the driving forces of actin-myosin binding. Cellular and Molecular Bioengineering. 6 (1), 26-37 (2013).
- Harada, Y., Sakurada, K., Aoki, T., Thomas, D. D., Yanagida, T. Mechanochemical coupling in actomyosin energy transduction by in vitro movement assay. Journal of Molecular Biology. 216 (1), 49-68 (1990).
- Fordyce, P. M., Valentine, M. T., Block, S. M. Advances in surface-based assays for single molecules. Single-Molecule Techniques: A Laboratory Manual. , 431-460 (2008).
- Ozeki, T., et al. Surface-bound casein modulates the adsorption and activity of kinesin on SiO2 surfaces. Biophysical Journal. 96 (8), 3305-3318 (2009).
- Neuman, K. C., Nagy, A. Single-molecule force spectroscopy: Optical tweezers, magnetic tweezers and atomic force microscopy. Nature Methods. 5 (6), 491-505 (2008).
- Neuman, K. C., Block, S. M.
- Thoresen, T., Lenz, M., Gardel, M. L. Thick filament length and isoform composition determine self-organized contractile units in actomyosin bundles. Biophysical Journal. 104 (3), 655-665 (2013).
- Matusovsky, O. S., et al. Millisecond conformational dynamics of skeletal Myosin II power stroke studied by high-speed atomic force microscopy. ACS Nano. 15 (2), 2229-2239 (2021).
- Reinemann, D. N., et al. Collective force regulation in anti-parallel microtubule gliding by dimeric Kif15 kinesin motors. Current Biology. 27 (18), 2810-2820 (2017).
- Cordova, J. C., et al. Bioconjugated core-shell microparticles for high-force optical trapping. Particle and Particle Systems Characterization. 35 (3), 1-8 (2018).
- Reinemann, D. N., Norris, S. R., Ohi, R., Lang, M. J. Processive Kinesin-14 HSET exhibits directional flexibility depending on motor traffic. Current Biology. 28 (14), 2356-2362 (2018).
- Forth, S., Hsia, K. C., Shimamoto, Y., Kapoor, T. M. Asymmetric friction of nonmotor MAPs can lead to their directional motion in active microtubule networks. Cell. 157 (2), 420-432 (2014).
- Shimamoto, Y., Kapoor, T. M. Analyzing the micromechanics of the cell division apparatus. Methods in Cell Biology. 145, 173-190 (2018).
- Shimamoto, Y., Forth, S., Kapoor, T. M. Measuring pushing and braking forces generated by ensembles of Kinesin-5 crosslinking two microtubules. Developmental Cell. 34 (6), 669-681 (2015).
- Thoresen, T., Lenz, M., Gardel, M. L.
- Murrell, M., Thoresen, T., Gardel, M.
- Weirich, K. L., Stam, S., Munro, E., Gardel, M. L. Actin bundle architecture and mechanics regulate myosin II force generation. Biophysical Journal. 120 (10), 1957-1970 (2021).
