

Summary
פעילות עצבית במערכות לעתים קרובות מחייב הפרשות סינכרוני פוטנציאל פעולה מתא עצב בתוך אוכלוסייה מסוימת. לדוגמה, פולסים של הורמון גונדוטרופין משחררים (GnRH) סביר לדרוש פעילות מתואמת בין הנוירונים GnRH. אנו מציגים גישה מתודולוגית שלנו אמין קבלת הקלטות אלקטרו סימולטני של נוירונים GnRH מופץ diffusely.
Abstract
גונדוטרופין, הורמון משחרר (GnRH) הוא neuropeptide קטן המווסת את בלוטת יותרת המוח לשחרר את הורמון luteinizing (LH) הורמון מגרה זקיק (FSH). Gonadotropins אלו הם חיוניים להסדרת פונקציית הרבייה. GnRH המכילים הנוירונים מופצים ברחבי diffusely ההיפותלמוס פרויקט לגדולה החציוני שם שחרור GnRH מ מסופי האקסון שלהם לתוך מערכת הפורטל hypophysiotropic (1). ב הנימים הפורטל, GnRH נוסע בלוטת יותרת המוח הקדמית כדי לעורר שחרור של gonadotropins למחזור מערכתית. שחרור GnRH אינו רציף אלא מופיע פולסים אפיזודי. היא מבוססת היטב כי בצורה מקוטעת של שחרור GnRH חיונית רבייה (2, 3).
תיאום פעילות של נוירונים GnRH מרובים כנראה בבסיס פולסים GnRH. תוכן הפפטיד GnRH סך נוירונים הוא כ 1.0 pg / התא (4), מתוכם 30% סביר כוללת את בריכת releasable. רמות של GnRH במהלך דופק (5, 6), מציע נוירונים GnRH מספר מעורבים כנראה neurosecretion. כמו כן, פעילות יחידה אחת מופק רב יחידת הקלטות ההיפותלמוס במהלך LH לשחרר מציין שינויים בפעילות של נוירונים מרובים (7). אלקטרודות עם פעילות שנרשמו במהלך פולסים LH המשויכים somata או GnRH או סיבים (8). לכן, לפחות חלק בפעילות זו נובעת נוירונים GnRH.
מנגנונים כי התוצאה ירי מסונכרן בנוירונים GnRH ההיפותלמוס אינם ידועים. הבהרת המנגנונים לתאם הירי בנוירונים GnRH היא בעיה מורכבת. ראשית, הנוירונים GnRH הם מעטים יחסית במספר. במכרסמים, יש 800-2500 נוירונים GnRH. אין זה ברור כי כל הנוירונים GnRH מעורבים שחרור GnRH אפיזודי. יתר על כן, הנוירונים GnRH מופצים diffusely (1). זה מסובך הבנתנו תיאום הירי עשה גישות טכניים רבים סורר. יש לנו אופטימיזציה רופף תאים המצורפת הקלטות במצב מהדק הנוכחי לאיתור ישיר של הפוטנציאלים לפעולה פיתחו גישה הקלטה המאפשר הקלטה בו זמנית של זוגות נוירונים GnRH.
Protocol
- פרוסות המוח ההיפותלמוס מוכנים, מודגרות והועבר לתא הקלטה כמפורט לעיל (9) אצל בעלי חיים אשר נוירונים GnRH לבטא חלבון פלואורסצנטי ירוק (GFP) בשליטת היזם את הפפטיד GnRH (10). לאחר הסרת המוח של החיה, המוח חסומה באמצעות סכין גילוח כדי למנוע אזורים שאינם בעלי עניין. על מנת לקבוע את האזורים להסיר, המוח הוא מונח על משטח הגב שלה, כך ההיפותלמוס ניתן דמיינו. במשך שתי אוריינטציות פרוסה העטרה ועל sagittal, במוח הקטן מוסר. להכנת פרוסה העטרה, חלק מקורי של קליפת מוסר. בכיוון זה פרוסה, האזורים לרוחב של קליפת לתמוך חותך את המוח לספק שטח מספיק של המוח עבור מיקום של חוטי כסף כדי להבטיח את המוח בחדר ההקלטה. להכנת פרוסה sagittal, אזורים הלטרלי של המוח יוסרו החלקים מקורי של קליפת לספק את התמיכה עבור המוח חיתוך ועל המיקום של חוטי כסף כדי להבטיח את המוח ההקלטה היטב. לאורך כל ההליך חוסם, המוח הוא ברציפות טבולה בנוזל השדרתי קר מלאכותי (ACSF) באמצעות פיפטה מזכוכית.
- כמות קטנה של הפלא מושם על מצע החיתוך. המוח הוא הרים במרית. ACSF עודף מוסר על ידי Kimwipe להביא אל פני השטח של המרית וציור ACSF עודף מן המוח. המוח הוא החליק בעדינות קדימה על הדבק. חשוב לא לגעת במוח או לדחוף אותו כלפי מטה על מנת לאבטח אותו. משקל המוח עצמו הוא בדרך כלל מספיק כדי להבטיח שהיא מאובטחת.
- המוח ואת פלטפורמת חיתוך מאובטח אז microtome הרוטט ואת חיתוך טוב מלא ACSF קר. הפתרון קר הן חברות המוח עשוי להגן מפני נזק במהלך הליך החיתוך. אזורי מוח שונים נבדלים במידת קור אשר מייעל פרוסות איכות. הטמפרטורה של ACSF להכנת פרוסות ההיפותלמוס הוא בדרך כלל 0 ° C.
- Slices צריך לחתוך לאט כל פרוסה צריך לצוף בחופשיות מן המוח. אם פרוסות מתחילים להסתלסל במהלך חיתוך, המהירות שבה הלהב מתקדמת יש האטה. ניתן להשתמש במברשת צבע קטנה פרוסות זזה אבל זה עושה את הסיכון לפגוע פרוסות ולכן, יש להימנע. לאט לאט למהירות מראש של הלהב הוא מועדף על פני שימוש במברשת צבע.
- כמו כל פרוסה חותכים מהמוח, היא נסוגה מן החדר חיתוך והניח בתא פרוסה הדגירה באמבט מים חמים (כ 32 ° C). פרוסה האינקובטור מסופק ברציפות עם חמצן באמצעות תערובת של 95% O 2 ו - 5% CO 2. בדרך כלל, מראש הקלטה הדגירה פרוסה נע בין 30 דקות עד 2 שעות.
- זכוכית pipettes (3-6 MΩ) מיוצרים כפי שתואר לעיל (9) באמצעות חולץ אנכי. הטמפרטורה ומשך משיכה משתנה בהתאם לסוג פיפטה חולץ אדם משתמש, סוג של כוס אחת משתמשת ואת חייו של נימה החימום של חולץ. צורת קצה פיפטה יכול גם להשתנות בהתאם להעדפה של המשתמש. השגנו הצלחה טוב עם קצה קוני אחיד עם פתח של 0.1 מ"מ. פתחים גדולים בתוך פיפטה נוטים נזק הנוירונים בזמן פתחים קטנים מצליחים לגרום הקלטות משך זמן רב.
- Pipettes מצופה Sylgard להפחית קיבול. טיפים פיפטה משך הם מצופים קלות Sylgard 184 באמצעות פיפטה השני. טיפים Sylgarded אז הם חום לרפא באמצעות אקדח חום. Pipettes אלה מתמלאים אז עם perfusate האמבטיה. כדי להגדיל את להדמיה של טיפים פיפטה, כמות קטנה של אלקסה-568 הוא הוסיף לחלק מהפתרון אמבטיה לשמש כפתרון פיפטה לפני מילוי pipettes. אנחנו לא שוקלים את אלקסה-568 אלא להשתמש מספיק כדי להפוך את ורוד פתרון על ידי העין. טיפים פיפטה לאחר מכן ניתן דמיינו תחת אדום טקסס הסינון.
- פיתחנו גישה להשיג מהדק השוטף של הקלטות עבור איתור של פוטנציאל פעולה באמצעות חותמות התנגדות נמוכה (ראה דיון). הקלטות מתבצעות על פוטנציאל המנוחה אנדוגני של התא עם Axoclamp 2B אקסון לא מיושם כיום כלי של משתמש. עם זאת, בשל לאטום את התנגדות נמוכה, לא ניתן היה לזהות פוטנציאל פעולה עם המגבר רק 2B. כדי להגביר את האות, מגבר שני (AM מערכות 3000) משמש בסדרת עם 2B Axoclamp. גישה זו יש את הקלות של חותמות רופף השירות לטווח ארוך הקלטות. מבחינה טכנית, אפשר למדוד ישירות הפוטנציאלים לפעולה, ובכך לעקוף את הבעיות עם הגישה במצב מתח, מהדק את ההקלטה (ראה דיון).
- אחד לוקח שני pipettes אל פני השטח של שני נוירונים שנבחרו קודם לכן באותו זמן, באופן דומה כמו הגישה עבור תצורת תא שלם ההקלטה. שניהם מתקרבים neurons באותו הזמן הצליחה יותר להשיג את הקלטות כפול מאשר לנסות כל נוירון של הצמד באופן עצמאי. במהלך תקופה זו, פולסים הזרקה הנוכחיים הם עברו המגבר 2B כפי שנעשה במהלך הגישה עבור כל תא הקלטות.
- לחץ חיובי יש לשמור על pipettes הן כדי למנוע סתימת קצה. לחץ חיובי עבור הקלטות כפול מופק הטובה ביותר באמצעות מזרק 3 מ"ל ללא מילוי ריק מצורף לבעל פיפטה לפי אורך של חתיכת צינור. באמצעות מזרקים מאפשר לשמור על הלחץ על pipettes שניהם. לחץ חיובי צריך להיות מיושם ברגע פיפטה הושם בפתרון באמבטיה של החדר ההקלטה. אחת הגישות הנוירונים נבחרת עם pipettes ברצף. לאחר מיצוב פיפטה first ישירות על פני השטח הגדול ביותר של הנוירון, אחד שומר חיובית על ידי השארת את המזרק במקום והעלים פיפטה בעמדה זו. אחת מכן מחזיר לחלק העליון של חדר הקלטה מוריד את פיפטה הקלטה השני למזיגה. נוירונים נבחרים לעיתים קרובות ברמות שונות של הפרוסה. פיפטה הראשון צריך להיות ממוקם מעל הנוירון כי הוא העמוק ביותר הפרוסה ואת עמדת פיפטה השני הנוירון שטחית יותר. זה מונע את המיקום של פיפטה השני הסטה את המיקום של פיפטה השני כמו פרוסת ינוע מעט במישור האנכי כמו pipettes להיכנס פרוסה.
- חותמות Loose (18-30 MΩ) ישמש. לחץ חיובי יניעו גומה קטנה על פני השטח של כל תא. אחת מנסה לאטום את נוירון לראשונה על ידי שחרור לחץ חיובי הפעלת לחץ שלילי קל מאוד. הלחץ החיובי הוא שוחרר על ידי הסרת מזרק. יניקה דרך הפה מוחל אז עד הסוף ללא צינורות. קרום תא עצב בריא יהיה להגיב במהירות כדי לשחרר את הלחץ החיובי לדבוק פיפטה. יניקה קלה יהוו חותמת עם התנגדות מתאימה נוירון בריא.
- כאשר אחד מנסה חותם נוירון ונכשל, פיפטה לא ניתן שימוש חוזר מאז ממברנות רק חותם על זכוכית נקי מאוד. במקום אחד מעלה את פיפטה (ואובייקטיבי מיקרוסקופ) מהמשטח של הפרוסה לראש זלוף הבאר, מסיר את פיפטה מהאמבטיה, שינויים פיפטה נקייה ואז חוזר הפרוסה לנסות לאטום תא אחר . מסיבה אלה, חשוב לבחור נוירונים פוטנציאל מרובים עבור הקלטות. עבור פרוסות עם פחות מ 4-6 איכות הקלטה נוירונים לבחירה, ההצלחה בהשגת הקלטות כפול מוגבל.
- לאחר הקמת חותם רופף על בסיס ההתנגדות, קטניות הזריקה הנוכחית מן המגבר 2B הן הסתיים. Amplifer 2B ממוקם סדרה עם amplifer 3000 הסדרה. בתצורה זו, את הפלט של amplifer 2B משמש כקלט עבור amplifer 3000 הסדרה. Amplifer האחרון מספק אות לרכישת נתונים.
- החותם (שלב 4) ו תצורה של amplifers (שלב 5) חוזר על עצמו ואז הנוירון השני.

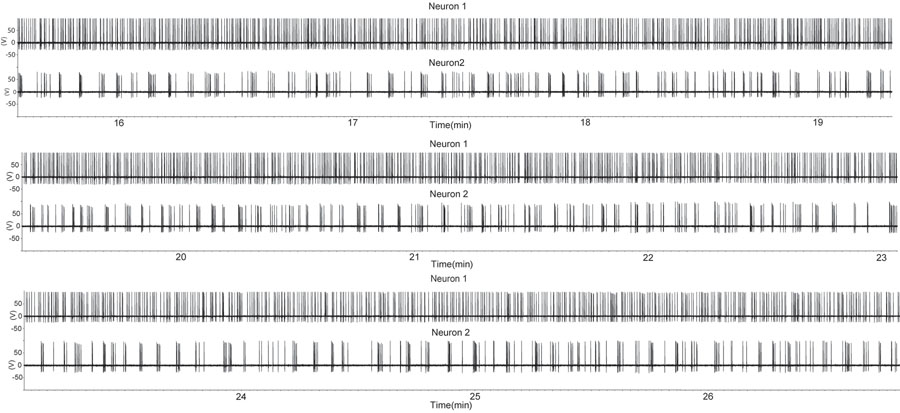
באיור 1. תקופה של עלייה בירי בשני נוירונים GnRH באמצעות חותם רופף מצורף תצורת הקלטה בהכנת פרוסה sagittal. פרוסה נגזר זכר מסורס. כל סטיה כלפי מעלה מצביע על פוטנציאל פעולה. שים לב כי אחד GnRH נוירון (עקבות למעלה) תערוכות ירי מתמשך כמעט ואילו השני GnRH נוירון תערוכות פרצי. דפוס זה של פעילות GnRH מתא עצב אחד דומה לזו של יחידות חילוץ מיחידת יחיד מרובה הקלטות במהלך הפרשת הורמון in vivo (7). אנא לחץ כאן כדי לראות גרסה גדולה יותר של דמות 1.
Discussion
הפעילות של עניין כמה נוירונים כולל נוירונים GnRH (מבוסס על הפרשת הורמון) מתרחשת על סולמות זמן של שעות (5-7). לכן, תצורת התא כולו אינה הבחירה הטובה ביותר עבור כמה מטרות הניסוי בשל דיאליזה של שליחים תאיים על מצב התא כולו ההקלטה. מטרה נוספת של כל תא הקלטות מוגבלות בדרך כלל בעלי חיים על פחות מ 120 ימים של גיל. עם הגיל, ממברנות עצביות להופיע להתקשות, מה שהופך חותמות עמידות גבוהה קשה להשיג. בנוסף, אם אחד מקבל חותם עמידות גבוהה, פקיעת התיקון משבש את החותם, והשאיר חור בין פיפטה לבין הממברנה. זה מוביל הקלטה שמיש ו נוירון כי ימות בתוך זמן קצר בשל חוסר האיזון היוני. מחזורי ביוץ רגיל, ולכן הפעילות יציבה של הגנרטור הדופק GnRH אינה מתרחשת עד מאוחר יותר בחיים (7-10 חודשים גיל C57Bl6 נקבות, 11, 12), מעבר לגיל שבו אפשר לצפות באופן סביר להשגת כל תא הקלטות אמין. לבסוף, כל תא הקלטות להרוס את יחסי אנדוגני של ריכוזי יון פנימיים וחיצוניים. עם הקלטות התא כולו, את הריכוז הפנימי של כל יון שווה את ריכוז היונים בתמיסה פיפטה. הסיבה לכך היא פתרון פיפטה של נפח גדול יחסית במהירות מגיע לשיווי משקל עם / מחליפה את אנדוגני נפח קטן יחסית של התא.
הגישה לתא רופף המצורף עוקף רבים המגבלות של כל תא הקלטות. ראשית, חותם התנגדות נמוכה (15-30 MΩ) יכול לשמש. אלה הם קלים יחסית טופס גם נוירונים מבעלי חיים מבוגרים. שנית, אחד לא קרע את התיקון של קרום אטום. לכן, תא הקלטות רופף מצורף מבחינה טכנית הרבה יותר קל מאשר כל תא הקלטות. בנוסף, מאז קרום התא לא נפגע, דיאליזה של רכיבים תאיים אינה מתרחשת ויחסים יוניים אנדוגני נשמרים. אי אפשר להשתמש בגישה תא רופף מצורף ללימוד זרמים סינפטיים אבל הוא אידיאלי עבור לטווח ארוך הקלטות מתא עצב בצורה יחסית לא פולשנית. תאים המצורפת הקלטות גם יכול להתבצע באמצעות כל פתרון תאיים סטנדרטי פיפטה. זה מציע יתרון נוסף של פקיעת תיקון קרום כאשר הקלטה ארוך טווח הושלם תיוג נוירון עם סמן תאיים.
הגישה לתא רופף מצורף נעשה שימוש במצב מתח, מהדק את ההקלטה. עם זאת, מתח-clamp הקלטה בתצורת תא רופף המצורפת יש בעיות מתודולוגיות שונות. ראשית, את האות מוקלט הוא אמצעי עקיף של פעילות. האות כי נמדד (כמו הפעולה שנקרא הנוכחי) הוא זרם קיבולי כי ההאשמות קרום (13). זוהי בעיה מתודולוגית חשובה ביותר. קיבול והתנגדות של טפטפת ההקלטה ניתן לסנן את האות מוקלט. ייתכן מאוד כי פעולה זרמים קטנים הם איבדו לחייב את הקיבול של פיפטה, אשר לא יכול להיות מתוגמל כראוי עם רוב מגברים, בשל התנגדות גבוהה של headstage. כאשר האותות הללו ללכת מבלי שיבחינו, דפוס ירי לכאורה של הנוירון אינו משקף את דפוס ירי אמיתי. כמו כן, פיפטה פיצוי ואת ההתנגדויות לגרום לשגיאות חותם משמעותי במהלך המדידות שינויים כגון כאשר זרמי פעולה באים לידי ביטוי (13). מגברים מסוימים לספק קיבול והתנגדות "פיצוי" על פיפטה ואת החותם, אשר מגביל אובדן האות, אך בראש מורם התנגדות שלבי ביותר מגברי לעכב פיצוי אופטימלי. שנית, מצב מלאכותי מוטל על התא. במצב מתח-clamp, האזור סביב קרום התא מוחזק הפוטנציאל קבוע, בכל המחקרים הללו, 0 mV. זה לא אומר שאין הנוכחית להחיל קרום התא. האות הנמדד מהדק מתח הוא למעשה כמות הזרם להחיל הממברנה כדי לשמור על פוטנציאל קבוע. לכן, הזרם הזה מיושם יכול לשנות את פעילות התא.
הקלטות כפול במערכת GnRH הם מאתגרת במיוחד בשל המספר המוגבל של נוירונים GnRH והפצה מפוזר שלהם. עבור הקלטות כפול כדי להצליח, מניפולטור חייב להיות יציב מאוד. אפילו תנועה קלה של האלקטרודה יכול לגרום פיפטה לחמוק מן הנוירון ולסיים את ההקלטה. יתר על כן, התנועה של פיפטה על התא (לדוגמא, מיצוב מחדש, כדי לפצות על התנועה) יכול לשנות את דפוסי הירי. כמה תעלות יונים כגון N-type ערוצי סידן רגישים מכנית: למתוח את הקרום גורמת פעילות חוזרת ונשנית בתצורות הקלטה הן כל תא ותא המצורפת (14). לבסוף, מערכת מניפולטור חייב להיות מסוגל תנועה יפה מאוד וחלק. כפי שצוין לעיל, עם הקלטות כפול, אחד לוקח שני pipettes אל פני השטח של שני נוירונים שנבחרו קודם לכן באותו זמן ומנסה לאטוםנוירון אחד. אם מצליחים, ואז אחד מנסה לאטום את התא השני. ככלל, לא ניתן לצפות לאטום ויש לי הקלטה באיכות גבוהה עם כל ניסיון. עם זאת, יוצרת בעיה מסוימת עם הקלטות כפול. אם הוא מוצלח עם הנוירון הראשון אך נכשל עם השני, יש לשנות את פיפטה ולנסות תא שונים. לכן, חייב להיות מסוגל להעביר גם את המטרה טבילה של המיקרוסקופ לבין פיפטה לראש זלוף היטב (כדי לשנות את פיפטה) מבלי לשבש את נוירון אטום בהצלחה.
הפיתוח שלנו ולהשתמש הגישה לתא מצורף רופף להקלטות כפול היא לקדם הטכניים העיקריים בלימוד נוירונים GnRH. זה עשוי להניב תוצאות שימושיות אשר יעזרו להזיז את השדה קדימה בהקשר של השאלה הקריטית של מה המנגנונים בבסיס פעילות מתואמת כי התוצאות הפרשת הורמון pulsatile.
Acknowledgments
אני אסיר תודה רונלד ל קלברזה, דיטר יגר (אוניברסיטת אמורי) וורד Yuhas (אקסון מכשירים) לדיונים טכניים שימושיים.
References
- Silverman, A. J. The gonadotropin releasing-hormone (GnRH) neuronal systems: immunocytochemistry. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Freeman, M. E. The neuroendocrine control of the ovarian cycle of the rat. In: The physiology of reproduction. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Belchetz, P. E., Plant, T. M., Nakai, Y., Keogh, E. J., Knobil, E. Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science. 202, 631-633 (1978).
- Maurer, J. A., Wray, S. Luteinizing hormone-releasing hormone quantified in tissues and slice explant cultures of postnatal rat hypothalami. Endocrinology. 140, 791-799 (1999).
- Harris, G. C., Levine, J. E. Pubertal acceleration of pulsatile gonadotropin-releasing hormone release in male rats as revealed by microdialysis. Endocrinology. 14, 163-171 (2003).
- Sisk, C. L., Richardson, H. N., Chappell, P. E., Levine, J. E. In vivo gonadotropin-releasing hormone secretion in female rats during peripubertal development and on proestrus. Endocrinology. 142, 2929-2936 (2001).
- Cardenas, H., Ordog, T., O'Byrne, K. T., Knobil, E. Single unit components of the hypothalamic multiunit electrical activity associated with the central signal generator that directs the pulsatile secretion of gonadotropic hormones. Proc Natl Acad Sci U S A. 90, 9630-9634 (1993).
- Silverman, A. J., Wilson, R., Kesner, J. S., Knobil, E. Hypothalamic localization of multiunit electrical activity associated with pulsatile LH release in the rhesus monkey. Neuroendocrinology. 44, 168-171 (1986).
- Roberts, C. B., O'Boyle, M. P., Suter, K. J. Dendrites determine the contribution of after depolarization potentials (ADPs) to generation of repetitive action potentials in hypothalamic gonadotropin releasing-hormone (GnRH) neurons. J Comput Neurosci. 26, 39-53 (2009).
- Spergel, D. J., Kruth, U., Hanley, D. F., Sprengel, R., Seeburg, P. H. GABA- and glutamate-activated channels in green fluorescent protein-tagged gonadotropin-releasing hormone neurons in transgenic mice. J Neurosci. 19, 2037-2050 (1999).
- Nelson, J. F., Felicio, L. S., Randall, P. K., Sims, C., Finch, C. E. A longitudinal study of estrous cyclicity in aging C57BL/6J mice: I. Cycle frequency, length and vaginal cytology. Biol Reprod. 27, 327-339 (1982).
- Gee, D. M., Flurkey, K., Finch, C. E. Aging and the regulation of luteinizing hormone in C57BL/6J mice: impaired elevations after ovariectomy and spontaneous elevations at advanced ages. Biol Reprod. 28, 598-607 (1983).
- Anson, B. D., Roberts, W. M. Loose-patch voltage-clamp technique. Neuromethods, Patch-clamp analysis: Advanced techniques. Walz, W., Boulton, A. A., Baker, G. B. 35, Humana Press. Place Unknown. 265-286 (2002).
- Calabrese, B., Tabarean, T., Juranka, P., Morris, C. E. Mechanosensitivity of N-type calcium channel currents. Biophys J. 83, 2560-2574 (2002).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Rapid killing of single neurons by irradiation of intracellularly injected dye. Science. 206, 702-704 (1979).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Lucifer Yellow slows voltage-gated Na+ current inactivation in a light-dependent manner in mice. J Physiol. 550, 159-167 (2003).

{kind=link}