

Summary
Activiteit in neuronale systemen vereist vaak synchroon actiepotentiaal lozingen afkomstig van neuronen in een specifieke populatie. Bijvoorbeeld, pulsen van gonadotrofine-releasing hormoon (GnRH) waarschijnlijk nodig gecoördineerde activiteit tussen GnRH neuronen. We presenteren onze methodische aanpak voor het verkrijgen van een betrouwbare gelijktijdige elektrofysiologische opnames van het diffuus verspreid GnRH neuronen.
Abstract
Gonadotrofine-releasing hormoon (GnRH) is een klein neuropeptide dat hypofyse afgifte van luteïniserend hormoon (LH) en follikelstimulerend hormoon (FSH) regelt. Deze gonadotropines zijn essentieel voor de regulering van de voortplantingsfunctie. De GnRH-bevattende neuronen zijn diffuus verspreid over het hele hypothalamus en project om de mediaan eminentie, waar ze vrij GnRH van hun axon terminals in het hypophysiotropic portal systeem (1). In het portaal haarvaten, GnRH reist naar de hypofyse tot afgifte van gonadotrofinen te stimuleren in de systemische circulatie. GnRH release is niet continu maar komt in episodische pulsen. Het staat vast dat de intermitterende wijze van GnRH release is van essentieel belang voor de voortplanting (2, 3).
Coördinatie van de activiteiten van meerdere GnRH neuronen waarschijnlijk ten grondslag ligt aan GnRH pulsen. Totaal peptide inhoud in GnRH neuronen is ongeveer 1,0 pg / cel (4), waarvan 30% bestaat uit de kans vrij te zwembad. Niveaus van GnRH tijdens een puls (5, 6), suggereren verschillende GnRH neuronen zijn waarschijnlijk betrokken bij neurosecretion. Ook enkele eenheid activiteit uit de hypothalamus multi-unit opnames tijdens de afgifte van het LH wijst op grote veranderingen in de activiteit van meerdere neuronen (7). De elektroden met geregistreerde activiteit tijdens LH pulsen worden geassocieerd met een GnRH somata of vezels (8). Daarom ten minste een aantal van deze activiteit komt voort uit GnRH neuronen.
De mechanismen die leiden tot een gesynchroniseerde bakken in de hypothalamus GnRH neuronen zijn onbekend. Ophelderen van de mechanismen die de coördinatie van vuren in GnRH neuronen is een complex probleem. Ten eerste, de GnRH neuronen zijn betrekkelijk gering in aantal. Bij knaagdieren, zijn er 800-2500 GnRH neuronen. Het is niet duidelijk dat alle GnRH neuronen betrokken zijn bij de episodische GnRH release. Bovendien worden GnRH neuronen diffuus verspreid (1). Dit heeft ingewikkelde ons begrip van de coördinatie van vuren en heeft tal van technische benaderingen hardnekkig. We hebben geoptimaliseerd losse cel-aangesloten opnames in de huidige-klem modus voor de directe detectie van actiepotentialen en ontwikkelde een opname aanpak die het mogelijk maakt om gelijktijdig opnamen van paren van GnRH neuronen.
Protocol
- Hypothalamus hersenen plakjes worden voorbereid, geïncubeerd en overgedragen aan de opname kamer zoals eerder gedetailleerd (9) in de dieren waarvan de GnRH neuronen te uiten groen fluorescerend eiwit (GFP) onder de controle van de GnRH peptide promotor (10). Na het verwijderen van de hersenen van het dier, is de hersenen geblokkeerd met behulp van een scheermesje om regio's die niet van belang zijn te elimineren. Om de regio's te verwijderen bepalen, wordt de hersenen gelegd op de dorsale oppervlak, zodat de hypothalamus kunnen worden gevisualiseerd. Voor zowel de coronale en sagittale slice oriëntaties, is het cerebellum verwijderd. Voor coronale slice voorbereiding, is de rostrale deel van de cortex verwijderd. In deze slice oriëntatie, de laterale gebieden van de cortex bieden ondersteuning voor het snijden van de hersenen en voldoende gebied van de hersenen voor de plaatsing van zilveren draden aan de hersenen in de opname kamer veilig te stellen. Voor de sagittale slice voorbereiding, zijn de laterale delen van de hersenen verwijderd en de rostrale delen van de cortex bieden de ondersteuning van de hersenen snijden en voor de plaatsing van zilver draden naar de hersenen in de opname goed te beveiligen. Gedurende de procedure te blokkeren, is het brein continu bevochtigd met koude kunstmatige cerebrospinale vloeistof (ACSF) met behulp van een glazen pipet.
- Een kleine hoeveelheid Superglue wordt geplaatst op de snij-platform. De hersenen zijn opgeheven met een spatel. Overtollig ACSF wordt verwijderd door te brengen een Kimwipe aan het oppervlak van de spatel en tekening overtollige ACSF uit de buurt van de hersenen. De hersenen zijn zacht naar voren geschoven op de lijm. Het is belangrijk om niet naar de hersenen raken of beneden duwt hem in om het te beveiligen. Het gewicht van de hersenen zelf is meestal voldoende om ervoor te zorgen dat het veilig is.
- De hersenen en het snijden platform is vervolgens aan de vibrerende microtoom en het snijden goed is gevuld met koud ACSF. De koude oplossing beide bedrijven de hersenen en kan tegen beschadiging te beschermen tijdens het snijden procedure. Verschillende hersengebieden variëren in de mate van kou die kwaliteit schijfjes optimaliseert. De temperatuur van de ACSF voor de voorbereiding van de hypothalamus plakken is over het algemeen 0 ° C.
- Slices moet langzaam worden gesneden en elk plakje moet vrij kunnen wegdrijven van de hersenen. Als plakjes beginnen te krullen tijdens het snijden, moet de snelheid waarmee het mes is oprukkende worden vertraagd. Men kan gebruik maken van een kleine kwast om uit de krul gaan plakken, maar dit betekent risico beschadiging van de plakjes en dus moet worden vermeden. Langzaam de snelheid van vooruitgang van het blad heeft de voorkeur boven het gebruik van een penseel.
- Omdat elk plakje wordt gesneden uit het brein, wordt het uit de snij-kamer en geplaatst in een plakje incubatiekamer in een warm waterbad (ongeveer 32 ° C). De slice incubator wordt continu voorzien van zuurstof met behulp van een mengsel van 95% O 2 en 5% CO 2. Over het algemeen, pre-opname slice incubatie varieert van 30 minuten tot 2 uur.
- Glazen pipetten (3-6 MΩ) zijn vervaardigd, zoals eerder beschreven (9) met behulp van een verticaal trekker. De temperatuur en de duur van het trekken is afhankelijk van de aard van de pipet trekker gebruikt men, het soort glas gebruikt men en de levensduur van de verwarming gloeidraad van de trekker. De vorm van de pipetpunt kan ook variëren afhankelijk van de voorkeur van de gebruiker. We hebben bereikt goed succes met een gelijkmatig conische tip met een opening van 0,1 mm. Grotere openingen in de pipet de neiging om de neuronen beschadigen terwijl de kleinere openingen niet te resulteren in langdurige opnames.
- Pipetten bedekt met Sylgard om capaciteit te verminderen. Getrokken pipetpunten zijn licht bedekt met Sylgard 184 met een tweede pipet. Sylgarded tips zijn dan warmte-cured met behulp van een warmte kanon. Deze pipetten worden vervolgens gevuld met het bad perfusaat. Ter verhoging van visualisatie van de pipet tips, is een kleine hoeveelheid van de Alexa-568 toegevoegd aan het gedeelte van het bad oplossing worden gebruikt als de pipet oplossing voor het vullen van de pipetten. Wij hebben geen wegen de Alexa-568, maar genoeg te gebruiken om de oplossing roze beurt door oog. De pipet tips kan dan zichtbaar worden onder de Texas rode filter.
- We hebben een aanpak ontwikkeld om de huidige-clamp opnames te verkrijgen voor de detectie van actiepotentialen met lage weerstand afdichtingen (zie Discussie). Opnames worden uitgevoerd op endogene van de cel rustpotentiaal met 2B zonder toegepaste stroom met behulp van Axon instrument Axoclamp. Echter, door de lage weerstand zegel, kon men niet detecteren actiepotentialen met alleen de 2B versterker. Voor het verhogen van het signaal, is een tweede versterker (AM Systems 3000) gebruikt in serie met de Axoclamp 2B. Deze aanpak heeft het gemak van losse zegels en het nut van de lange termijn opnames. Technisch gezien kan men direct te meten actiepotentialen, waardoor het omzeilen van de problemen met de aanpak in de voltage-clamp opnamemodus (zie Discussie).
- Men neemt twee pipetten aan het oppervlak van de twee eerder geselecteerde neuronen op hetzelfde moment, op een soortgelijke manier als de aanpak voor de hele cel opname configuratie. Het naderen van beide neurons tegelijkertijd is meer succes in het verkrijgen van de dubbele opnames van te proberen elk neuron van het paar onafhankelijk van elkaar. Gedurende deze tijd zijn de huidige injectie impulsen doorgegeven van de 2B versterker zoals dat ook gebeurt bij de aanpak voor whole-cell opnames.
- Positieve druk moet worden gehandhaafd op beide pipetten om te voorkomen dat verstopping van de tip. Positieve druk voor dual-opnames is het best gegenereerd door het gebruik van een ongevulde 3 ml lege spuit aan de pipet houder door een lengte van stuk buis. Met behulp van spuiten maakt het mogelijk om te onderhouden de druk op beide pipetten. Positieve druk dient zo snel als de pipet is geplaatst in het bad oplossing van de opname kamer worden toegepast. Een aanpak van de geselecteerde neuronen met de pipetten opeenvolgend. Na het plaatsen van de eerste pipet direct over het grootste oppervlak van het neuron, handhaaft een positieve Bij het verlaten van de spuit op zijn plaats en laat de pipet in deze positie. Men keert terug naar de bovenkant van de opname kamer en verlaagt de tweede opname pipet in de juiste positie. Geselecteerde neuronen worden vaak op verschillende niveaus van de slice. De eerste pipet moet worden geplaatst op het neuron dat diepste in de slice en de positie van de tweede pipet op de meer oppervlakkige neuron. Dit voorkomt dat de positionering van de tweede pipet uit het verschuiven van de positionering van de tweede pipet als de slice zal iets bewegen in het verticale vlak als pipetten voert u het schijfje.
- Losse zegels (18-30 MΩ) zal worden gebruikt. De positieve druk zal leiden tot een klein kuiltje op het oppervlak van elke cel. Men probeert om de eerste neuron zegel door het vrijgeven van positieve druk en het toepassen van zeer lichte onderdruk. De positieve druk wordt vrijgegeven door het verwijderen van de spuit. Afzuiging via de mond wordt vervolgens toegepast op het vrije uiteinde van de slang. Een gezonde neuron membraan zal snel reageren op de vrijlating van positieve druk en zich houden aan de pipet. Een geringe onderdruk vormt een afdichting met de juiste weerstand in een gezonde neuron.
- Wanneer men probeert te verzegelen een neuron en faalt, kan de pipet niet opnieuw worden gebruikt, omdat membranen alleen afdichting tot zeer schoon glas. In plaats daarvan, een verhoogt de pipet (en microscoop objectief) uit de buurt van het oppervlak van de slice naar de top van de perfusie goed, verwijdert de pipet uit het bad, verandert in een schone pipet en vervolgens terugkeert naar de slice om te proberen naar een andere cel afdichting . Voor deze reden is het belangrijk om u meerdere mogelijkheden neuronen voor opnamen. Voor de plakjes met minder dan 4-6 opnamekwaliteit neuronen uit te kiezen, het succes bij het verkrijgen van dual-opnames is beperkt.
- Na de oprichting van een losse afdichting op basis van weerstand, zijn de huidige injectie pulsen van de 2B versterker beëindigd. Het 2B versterker is geplaatst in serie met de 3000-serie versterker. In deze configuratie, de output van de 2B versterker dient als input voor de 3000-serie versterker. De laatste versterker biedt signaal voor de data-acquisitie.
- De afdichting (stap 4) en herconfiguratie van amplifers (stap 5) wordt vervolgens herhaald voor de tweede neuron.

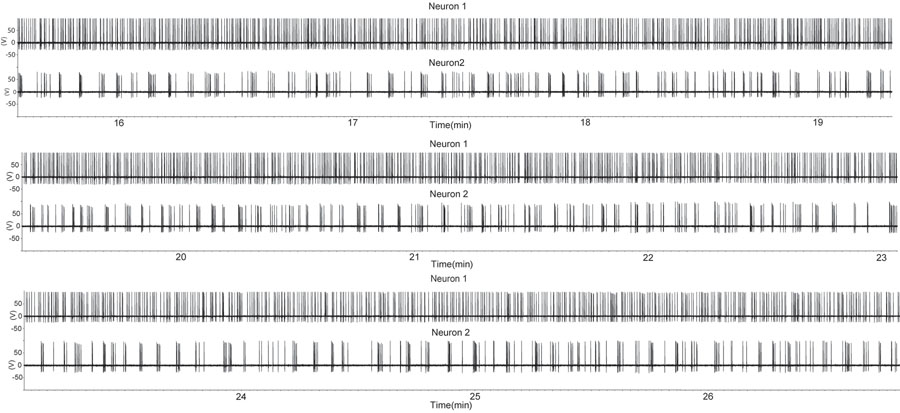
Figuur 1. Een periode van verhoogde vuren in twee GnRH neuronen met behulp van de losse zegel bevestigd opname configuratie in een sagittale slice voorbereiding. De slice is afgeleid van een gecastreerde reu. Elke opwaartse doorbuiging geeft een actiepotentiaal. Merk op dat een GnRH neuron (top sporen) bijna continu branden vertoont, terwijl de tweede GnRH neuron vertoont intermitterende barst. Dit patroon van activiteit van een GnRH-neuronen is vergelijkbaar met die van de afzonderlijke eenheden uit multi-unit opnames tijdens het hormoon in vivo (7). Gelieve Klik hier voor een grotere versie van figuur 1 te zien.
Discussion
De activiteit van belang in sommige neuronen met GnRH-neuronen (gebaseerd op de hormoon) komt op een tijdschaal van uren (5-7). Daarom is de hele cel configuratie is niet de beste keuze voor een aantal experimentele doelen als gevolg van de dialyse van intracellulaire boodschappers in de hele cel opnamemodus. Secundair, zijn hele-cel-opnamen over het algemeen beperkt tot dieren van minder dan ongeveer 120 dagen oud. Met de leeftijd, de neuronale membranen lijken te verstijven, waardoor een hoge weerstand tegen zegels moeilijk te bereiken. Bovendien, als verkrijgt men een hoge weerstand zegel, scheuren de patch verstoort de afdichting, waardoor er een gat tussen de pipet en het membraan. Dit leidt tot een onbruikbare opname en een neuron die snel zullen sterven als gevolg van ionische onevenwichtigheden. Regelmatige ovariële cyclus, en dus, stabiele activiteit van de GnRH pulsgenerator doet zich niet pas later in het leven (7-10 maanden van leeftijd in C57Bl6 vrouwen, 11, 12), na de leeftijd waarop een redelijkerwijs kunnen anticiperen op het verkrijgen van whole-cell-opnamen betrouwbaar. Tot slot, whole-cell-opnamen te vernietigen de endogene ratio's van interne en externe ion concentraties. Met hele cel-opnamen, de interne concentratie van een ion is gelijk aan de concentratie van het ion in de pipet oplossing. Dit komt omdat de pipet oplossing is relatief groot volume snel in evenwicht is gekomen met / vervangt de relatief kleine endogene volume van de cel.
De losse cel bevestigd aanpak omzeilt veel van de beperkingen van whole-cell opnames. Ten eerste kan een lage weerstand afdichting (15-30 MΩ) worden gebruikt. Deze zijn relatief eenvoudig te vormen, zelfs in neuronen van oudere dieren. Ten tweede, men niet breken de verzegelde stukje membraan. Daarom, losse cel bevestigd opnames zijn technisch veel makkelijker dan whole-cell opnames. Bovendien, omdat de celmembraan intact is, is dialyse van intracellulaire componenten niet voorkomen en endogene ionische ratio's zijn bewaard gebleven. Men kan de losse cel bevestigd aanpak niet gebruiken voor het bestuderen van synaptische stromen, maar het is ideaal voor lange-termijn opnamen van neuronen in een relatief niet-invasieve manier. De cel-ingeschreven opnames kunnen ook worden uitgevoerd met behulp van een standaard intracellulaire oplossing in de pipet. Dit biedt het bijkomende voordeel van scheurt het membraan patch als de lange termijn opname is voltooid en de etikettering van de neuron met een intracellulaire marker.
De losse cel bevestigd benadering is gebruikt in de voltage-clamp opnamemodus. Echter, voltage-clamp opname in de losse cel bevestigd configuratie heeft een aantal methodologische problemen. Eerste, het opgenomen signaal is een indirecte meting van de activiteit. Het signaal dat wordt gemeten (zoals de zogenaamde actie stroom) is de capacitieve stroom die het membraan (13) kosten. Dit is een zeer belangrijke methodologische kwestie. De capaciteit en weerstand van een opname pipet kan filteren het opgenomen signaal. Het is zeer waarschijnlijk dat de kleine actie stromingen zijn verloren in het opladen van de capaciteit van de pipet, die niet goed kan worden gecompenseerd met de meeste versterkers, vanwege de hoge weerstand van de headstage. Wanneer deze signalen onopgemerkt blijven, is de schijnbare afvuren patroon van het neuron geen afspiegeling van de ware vuren patroon. Ook niet-gecompenseerde pipet en afdichting weerstanden leiden tot ernstige fouten in de metingen tijdens veranderingen, zoals bij de actie stromen worden uitgedrukt (13). Sommige versterkers geven de capaciteit en het verzet "compensatie" voor de pipet en afdichting, die signaalverlies beperkt, maar een hoge weerstand tegen het hoofd stadia van de meeste versterkers belemmeren optimale compensatie. Ten tweede wordt een kunstmatige situatie die aan de cel. In voltage-clamp-modus, is het gebied rondom de celmembraan gehouden om een vaste potentiaal, in deze studies, 0 mV. Dit betekent niet dat er geen stroom toegepast op de celmembraan. Het signaal gemeten in voltage-clamp is eigenlijk de hoeveelheid stroom die wordt toegepast om het membraan naar de vaste potentiaal te houden. Daarom kan deze toegepast stroom aan te passen van de cel-activiteit.
Dual opnamen in het GnRH-systeem zijn bijzonder uitdagend te wijten aan het beperkte aantal van GnRH neuronen en hun diffuse distributie. Voor dual-opnamen om succesvol te zijn, moet de manipulator worden buitengewoon stabiel. Zelfs lichte beweging van de elektrode kan ertoe leiden dat de pipet te glijden uit de neuron en de opname te beëindigen. Bovendien kan beweging van de pipet op de cel (bijvoorbeeld re-positionering te compenseren voor de beweging) te veranderen vuren patronen. Sommige ion kanalen zoals N-type calciumkanalen zijn mechanisch gevoelig: membraan stretch oorzaken repetitieve activiteit in beide whole-cel en cel-aangesloten opname configuraties (14). Ten slotte moet de manipulator systeem in staat zijn uiterst fijn en vloeiende bewegingen. Zoals hierboven vermeld, met dubbele opnames, neemt men twee pipetten aan het oppervlak van de twee eerder geselecteerde neuronen op hetzelfde moment en probeert af te dichteneen neuron. Als dat lukt, dan een poging om de tweede cel afdichting. In het algemeen kan men niet verwachten dat afdichting en een hoge kwaliteit opname te hebben met elke poging. Dit, echter, schept een bepaald probleem met dubbele opnamen. Als men succesvol is met het eerste neuron, maar niet in slaagt met de tweede, moet een verandering van de pipet en probeer een andere cel. Daarom moet men in staat zijn om zowel de immersie objectief van de microscoop en de pipet naar het begin van de perfusie goed (om de pipet te wijzigen) zonder verstoring van de succesvol afgesloten neuron.
Onze ontwikkeling en het gebruik van de losse cel-aangesloten aanpak voor dual-opnames is een belangrijke technische vooruitgang in het bestuderen van GnRH neuronen. Het is waarschijnlijk om nuttige resultaten die zullen helpen vooruit het veld in het kader van de kritische vraag wat de mechanismen ten grondslag liggen aan de gecoördineerde activiteit die resulteert in een pulsatiele secretie hormoon. Produceren
Acknowledgments
Ik ben blij dat Ronald L. Calabrese, Dieter Jaeger (Emory University) en Ward Yuhas (Axon Instruments) voor nuttige technische besprekingen.
References
- Silverman, A. J. The gonadotropin releasing-hormone (GnRH) neuronal systems: immunocytochemistry. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Freeman, M. E. The neuroendocrine control of the ovarian cycle of the rat. In: The physiology of reproduction. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Belchetz, P. E., Plant, T. M., Nakai, Y., Keogh, E. J., Knobil, E. Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science. 202, 631-633 (1978).
- Maurer, J. A., Wray, S. Luteinizing hormone-releasing hormone quantified in tissues and slice explant cultures of postnatal rat hypothalami. Endocrinology. 140, 791-799 (1999).
- Harris, G. C., Levine, J. E. Pubertal acceleration of pulsatile gonadotropin-releasing hormone release in male rats as revealed by microdialysis. Endocrinology. 14, 163-171 (2003).
- Sisk, C. L., Richardson, H. N., Chappell, P. E., Levine, J. E. In vivo gonadotropin-releasing hormone secretion in female rats during peripubertal development and on proestrus. Endocrinology. 142, 2929-2936 (2001).
- Cardenas, H., Ordog, T., O'Byrne, K. T., Knobil, E. Single unit components of the hypothalamic multiunit electrical activity associated with the central signal generator that directs the pulsatile secretion of gonadotropic hormones. Proc Natl Acad Sci U S A. 90, 9630-9634 (1993).
- Silverman, A. J., Wilson, R., Kesner, J. S., Knobil, E. Hypothalamic localization of multiunit electrical activity associated with pulsatile LH release in the rhesus monkey. Neuroendocrinology. 44, 168-171 (1986).
- Roberts, C. B., O'Boyle, M. P., Suter, K. J. Dendrites determine the contribution of after depolarization potentials (ADPs) to generation of repetitive action potentials in hypothalamic gonadotropin releasing-hormone (GnRH) neurons. J Comput Neurosci. 26, 39-53 (2009).
- Spergel, D. J., Kruth, U., Hanley, D. F., Sprengel, R., Seeburg, P. H. GABA- and glutamate-activated channels in green fluorescent protein-tagged gonadotropin-releasing hormone neurons in transgenic mice. J Neurosci. 19, 2037-2050 (1999).
- Nelson, J. F., Felicio, L. S., Randall, P. K., Sims, C., Finch, C. E. A longitudinal study of estrous cyclicity in aging C57BL/6J mice: I. Cycle frequency, length and vaginal cytology. Biol Reprod. 27, 327-339 (1982).
- Gee, D. M., Flurkey, K., Finch, C. E. Aging and the regulation of luteinizing hormone in C57BL/6J mice: impaired elevations after ovariectomy and spontaneous elevations at advanced ages. Biol Reprod. 28, 598-607 (1983).
- Anson, B. D., Roberts, W. M. Loose-patch voltage-clamp technique. Neuromethods, Patch-clamp analysis: Advanced techniques. Walz, W., Boulton, A. A., Baker, G. B. 35, Humana Press. Place Unknown. 265-286 (2002).
- Calabrese, B., Tabarean, T., Juranka, P., Morris, C. E. Mechanosensitivity of N-type calcium channel currents. Biophys J. 83, 2560-2574 (2002).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Rapid killing of single neurons by irradiation of intracellularly injected dye. Science. 206, 702-704 (1979).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Lucifer Yellow slows voltage-gated Na+ current inactivation in a light-dependent manner in mice. J Physiol. 550, 159-167 (2003).

{kind=link}