

Summary
L'attività in sistemi neuronali spesso richiede scariche azione sincrona potenziale da parte dei neuroni all'interno di una popolazione specifica. Per esempio, gli impulsi di gonadotropina-releasing hormone (GnRH) probabilmente bisogno di una attività coordinata tra i neuroni GnRH. Vi presentiamo il nostro approccio metodologico affidabile per ottenere simultanea registrazioni elettrofisiologiche da neuroni diffusamente distribuite GnRH.
Abstract
Gonadotropina-Releasing Hormone (GnRH) è un piccolo neuropeptide che regola liberazione ipofisaria di ormone luteinizzante (LH) e ormone follicolo-stimolante (FSH). Queste gonadotropine sono essenziali per la regolazione della funzione riproduttiva. Il GnRH contenenti neuroni sono distribuiti capillarmente in tutta l'ipotalamo e il progetto per l'eminenza mediana dove rilascio GnRH dai loro terminali degli assoni nel sistema portale hypophysiotropic (1). Nei capillari portale, GnRH viaggia alla ghiandola pituitaria anteriore di stimolare il rilascio di gonadotropine in circolazione sistemica. Rilascio del GnRH non è continua ma avviene invece in impulsi episodici. E 'ben noto che il modo intermittente del rilascio di GnRH è essenziale per la riproduzione (2, 3).
Coordinamento delle attività dei neuroni GnRH più probabilmente alla base di impulsi di GnRH. Peptide contenuto totale in neuroni GnRH è di circa 1,0 pg / cell (4), di cui il 30% comprende probabilmente la piscina rilasciabili. I livelli di GnRH durante un impulso (5, 6), suggeriscono più neuroni GnRH sono probabilmente coinvolti in neurosecretion. Allo stesso modo, l'attività singola unità estratto da ipotalamo multi-unità registrazioni durante LH rilascio indicano cambiamenti nell'attività dei neuroni più (7). Gli elettrodi con attività registrate durante gli impulsi LH sono associati sia con somata GnRH o fibre (8). Quindi, almeno alcune di queste attività nasce da neuroni GnRH.
I meccanismi che provocano la cottura sincronizzati nei neuroni GnRH ipotalamici sono sconosciuti. Chiarire i meccanismi che coordinano la cottura nei neuroni GnRH è un problema complesso. In primo luogo, i neuroni GnRH sono relativamente poco numerosi. Nei roditori, ci sono 800-2500 neuroni GnRH. Non è chiaro che tutti i neuroni GnRH sono coinvolti nel rilascio del GnRH episodica. Inoltre, i neuroni GnRH sono diffusamente distribuite (1). Questo ha complicato la nostra comprensione di coordinamento di cottura e ha fatto molti approcci tecnici intrattabile. Abbiamo ottimizzato perdere cellula-attached registrazioni in current-clamp modalità per la rilevazione diretta dei potenziali d'azione e ha sviluppato un metodo di registrazione che permette di registrazioni simultanee da coppie di neuroni GnRH.
Protocol
- Sezioni di cervello ipotalamici sono preparati, incubate e trasferiti alla camera di registrazione come precedentemente dettagliato (9) negli animali il cui neuroni GnRH esprimere una proteina fluorescente verde (GFP) sotto il controllo del promotore peptide GnRH (10). Dopo la rimozione del cervello dall'animale, il cervello è bloccato con una lama di rasoio per eliminare le regioni che non sono di interesse. Al fine di determinare le regioni di rimuovere, il cervello è posto sulla sua superficie dorsale in modo che l'ipotalamo possono essere visualizzati. Per entrambi gli orientamenti fetta coronale e sagittale, il cervelletto viene rimosso. Per la preparazione fetta coronale, la porzione rostrale della corteccia è stato rimosso. In questo orientamento fetta, le regioni della corteccia laterale forniscono il supporto per affettare il cervello e fornire spazio sufficiente di cervello per il posizionamento di fili d'argento per proteggere il cervello nella camera di registrazione. Per la preparazione fetta sagittale, le regioni laterali del cervello vengono rimossi e la parte rostrale della corteccia fornisce il supporto per il cervello affettare e per il posizionamento di fili d'argento per proteggere il cervello nella registrazione bene. Nel corso della procedura di blocco, il cervello è continuamente inumidito con il freddo liquido cerebrospinale artificiale (ACSF) usando una pipetta di vetro.
- Una piccola quantità di colla è posto sulla piattaforma di taglio. Il cervello viene sollevato con una spatola. ACSF eccesso viene rimosso da portare un Kimwipe alla superficie della spatola e il disegno ACSF in eccesso dal cervello. Il cervello è dolcemente scivolare in avanti sulla colla. E 'importante non toccare il cervello o spingere verso il basso su di esso per fissarlo. Il peso del cervello stesso è solitamente sufficiente per assicurarsi che sia sicuro.
- Il cervello e la piattaforma di taglio è poi fissato per il microtomo vibrante e il taglio bene è pieno di freddo ACSF. La soluzione fredda entrambe le aziende il cervello e può proteggere da eventuali danni durante la procedura di taglio. Differenti regioni del cervello variano nel grado di freddezza che ottimizza fette di qualità. La temperatura del ACSF per la preparazione di fette ipotalamo è generalmente 0 ° C.
- Fette devono essere tagliate ogni fetta lentamente e dovrebbe fluttuare liberamente lontano dal cervello. Se le slice cominciano ad arricciarsi durante il taglio, la velocità con cui avanza la lama deve essere rallentato. Si può utilizzare un piccolo pennello di vernice a fette uncurl ma questo non rischia di danneggiare le fette e, quindi, dovrebbe essere evitato. Lentamente, la velocità di avanzamento della lama è preferibile usare un pennello.
- Come ogni fetta è tagliato dal cervello, si è ritirato dalla camera di taglio e posto in una camera di incubazione fetta in un bagno di acqua calda (circa 32 ° C). La fetta incubatore è continuamente fornito con l'ossigeno con una miscela di 95% O 2 e il 5% di CO 2. In generale, pre-registrazione va incubazione fetta da 30 minuti a 2 ore.
- Pipette di vetro (3-6 MΩ) sono realizzati come descritto in precedenza (9) con un estrattore verticale. La temperatura e la durata della trazione varia in base al tipo di pipetta estrattore si usa, il tipo di vetro si usa e la durata del filamento di riscaldamento l'estrattore. La forma del puntale può anche variare a seconda delle preferenze dell'utente. Abbiamo raggiunto un buon successo con una punta uniformemente conica con un'apertura di 0,1 mm. Grandi aperture nella pipetta tendono a danneggiare i neuroni, mentre le aperture più piccole non riescono a portare a registrazioni di lunga durata.
- Pipette rivestito con Sylgard per ridurre la capacità. Puntali sono leggermente tirato rivestiti con Sylgard 184 con una pipetta secondo. Suggerimenti Sylgarded sono poi calore curato con una pistola di calore. Queste pipette vengono poi riempite con il perfusato bagno. Per aumentare la visualizzazione dei puntali, una piccola quantità di Alexa-568 si aggiunge alla parte della soluzione da bagno da utilizzare come soluzione pipetta prima di riempire le pipette. Noi non pesare i Alexa-568, ma piuttosto l'uso abbastanza per girare la rosa soluzione ad occhio. Il puntali possono poi essere visualizzate sotto il filtro rosso Texas.
- Abbiamo sviluppato un approccio per ottenere corrente-clamp registrazioni per l'individuazione di potenziali d'azione con guarnizioni a bassa resistenza (vedi Discussione). Le registrazioni vengono effettuate a potenziale endogeno di riposo della cellula con Axoclamp 2B non applicata attuale Strumento Axon utilizzando il. Tuttavia, a causa della tenuta bassa resistenza, non in grado di rilevare potenziali d'azione con il solo amplificatore 2B. Per aumentare il segnale, un secondo amplificatore (AM Sistemi 3000) viene utilizzato in serie con la 2B Axoclamp. Questo approccio ha la facilità di foche sciolti e l'utilità di lungo periodo delle registrazioni. Tecnicamente, si può misurare direttamente i potenziali di azione, aggirando in tal modo i problemi con l'approccio del voltage-clamp modalità di registrazione (vedi Discussione).
- Si prende due pipette alla superficie dei due neuroni selezionati in precedenza allo stesso tempo, in modo simile come approccio per l'intera configurazione di registrazione delle cellule. Avvicinandosi entrambi neurons allo stesso tempo è stato più riuscito ad ottenere le registrazioni duale che tentare ogni neurone della coppia in modo indipendente. Durante questo tempo, gli impulsi di iniezione di corrente sono passati da amplificatore 2B come si fa durante l'avvicinamento per intero a cellule registrazioni.
- Pressione positiva deve essere mantenuta da entrambe le pipette per evitare l'intasamento della punta. Pressione positiva per le registrazioni duale è meglio generata utilizzando un vuoto non riempito siringa da 3 ml legata al titolare pipettare con una lunghezza del pezzo di tubo. Utilizzando siringhe permette di mantenere la pressione su entrambe le pipette. Pressione positiva dovrebbe essere applicato non appena la pipetta è stata posta nella soluzione bagno della camera di registrazione. Ci si avvicina alla neuroni selezionati con le pipette in modo sequenziale. Dopo aver posizionato la pipetta prima direttamente sopra la più grande superficie del neurone, si mantiene positivo da parte lasciando la siringa in posizione e lascia la pipetta in questa posizione. Si ritorna poi alla cima della camera di registrazione e abbassa la pipetta secondi di registrazione in posizione. Neuroni selezionati sono spesso a diversi livelli della fetta. La pipetta primo dovrebbe essere posizionato sopra il neurone che è più profondo nella fetta e posizionare la pipetta secondo al neurone più superficiali. Questo impedisce il posizionamento della pipetta secondo dal spostando il posizionamento della pipetta secondo come la fetta si sposta leggermente sul piano verticale come pipette inserire la fetta.
- Guarnizioni allentate (18-30 MΩ) verrà utilizzato. La pressione positiva indurrà una piccola fossetta sulla superficie di ogni cellula. Un tentativo di sigillare il primo neurone rilasciando pressione positiva e applicando molto lieve pressione negativa. La pressione positiva viene rilasciato rimuovendo la siringa. Aspirazione per via orale viene poi applicato alla estremità libera del tubo. Membrana Un neurone sano che rispondere rapidamente al rilascio di pressione positiva e di aderire alla pipetta. Aspirazione leggera formerà un sigillo con resistenza appropriata in un neurone sano.
- Quando si tenta di sigillare un neurone e non riesce, la pipetta non può essere riutilizzato dal membrane solo sigillo per pulire molto vetro. Invece, si solleva la pipetta (e obiettivo microscopio) di distanza dalla superficie della fetta nella parte superiore della perfusione bene, toglie la pipetta dal bagno, si trasforma in una pipetta pulita e poi torna alla fetta per tentare di sigillare un'altra cella . Per queste ragioni, è importante per selezionare più neuroni potenziale per le registrazioni. Per le fette con neuroni di registrazione meno di 4-6 qualità tra cui scegliere, il successo ad ottenere le registrazioni doppio è limitata.
- Dopo aver stabilito un sigillo sciolto in base alla resistenza, gli impulsi di iniezione di corrente dall'amplificatore 2B sono terminati. L'amplificatore 2B è posto in serie con l'amplificatore serie 3000. In questa configurazione, l'uscita del amplificatore 2B serve come ingresso per l'amplificatore serie 3000. L'amplificatore Quest'ultimo fornisce il segnale per l'acquisizione dati.
- La tenuta (punto 4) e la riconfigurazione di amplificatori (punto 5) viene quindi ripetuto per il neurone secondo.

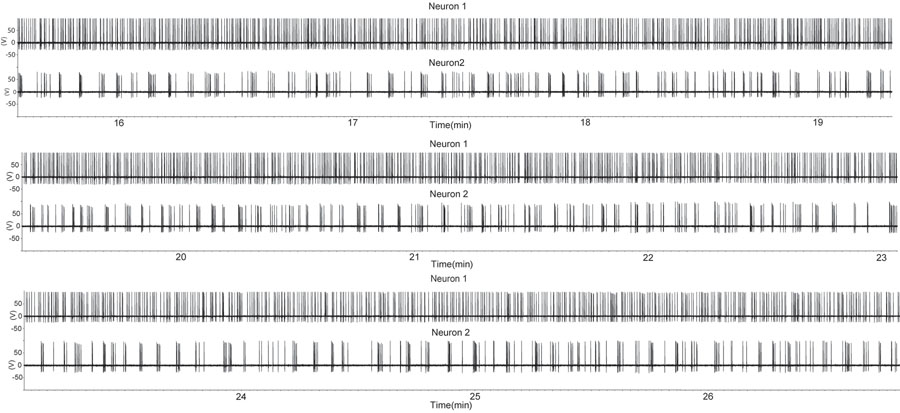
Figura 1. Un periodo di maggiore cottura in due neuroni a piede libero con GnRH configurazione di registrazione allegata sigillo in un preparato fetta sagittale. La fetta è stata derivata da un maschio castrato. Ogni deviazione verso l'alto indica un potenziale d'azione. Si noti che un neurone GnRH (traccia in alto) mostra cottura quasi continuo, mentre il secondo neurone GnRH presenta scoppi intermittenti. Questo modello di attività di singoli neuroni GnRH è simile a quella di singole unità estratte da più unità registrazioni durante la secrezione dell'ormone in vivo (7). Si prega di cliccare qui per vedere una versione più grande della figura 1.
Discussion
L'attività di interesse in alcuni neuroni compresi i neuroni GnRH (basato sulla secrezione ormonale) si verifica su scale di tempo di ore (5-7). Pertanto, la configurazione di cellula intera non è la scelta migliore per alcuni obiettivi sperimentali a causa della dialisi di messaggeri intracellulari in modalità intera registrazione delle cellule. Secondariamente, a cellula intera registrazioni sono generalmente limitata agli animali meno di circa 120 giorni di età. Con l'età, membrane neuronali sembrano irrigidirsi, rendendo guarnizioni ad alta resistenza difficile da raggiungere. Inoltre, se si ottiene un sigillo ad alta resistenza, rompendo la patch rompe il sigillo, lasciando un buco tra la pipetta e la membrana. Questo porta ad una registrazione inutilizzabile e un neurone che presto morirà a causa di squilibri ionici. Cicli ovarici normali e, quindi, l'attività stabile del generatore di impulsi GnRH non si verifica fino a quando nel corso della vita (7-10 mesi di età in C57Bl6 femmine, 11, 12), oltre l'età in cui si può ragionevolmente prevedere di ottenere cellule intere registrazioni affidabile. Infine, a cellula intera registrazioni distruggere i rapporti endogena di concentrazioni di ioni interne ed esterne. Con le registrazioni cellula intera, la concentrazione interna di ogni ione è uguale alla concentrazione dello ione nella soluzione pipetta. Questo perché il volume della soluzione di pipetta è relativamente grande raggiunge rapidamente l'equilibrio con / sostituisce il volume relativamente piccolo endogena della cellula.
L'approccio delle cellule perdere attaccato aggira molte delle limitazioni di cellule intere registrazioni. In primo luogo, una tenuta bassa resistenza (15-30 MΩ) può essere utilizzato. Queste sono relativamente facili da formare, anche nei neuroni di animali più vecchi. In secondo luogo, non si rompe la patch di membrana sigillata. Pertanto, le registrazioni delle cellule perdere allegato sono tecnicamente molto più facile rispetto a cellule intere registrazioni. Inoltre, poiché la membrana cellulare è intatto, la dialisi di componenti intracellulari non si verifica e endogeni rapporti ionici sono conservati. Non si può utilizzare l'approccio sciolto cellulare attaccato per studiare le correnti sinaptiche, ma è l'ideale per le registrazioni a lungo termine da parte dei neuroni in maniera relativamente non invasiva. La cellula-attached registrazioni possono essere eseguite anche utilizzando qualsiasi soluzione standard intracellulare nella pipetta. Questo offre l'ulteriore vantaggio di rottura della patch membrana quando la registrazione a lungo termine è completato e l'etichettatura del neurone con un marcatore intracellulare.
L'approccio delle cellule perdere allegato è stato utilizzato in voltage-clamp modalità di registrazione. Tuttavia, voltage-clamp registrazione nella configurazione della sciolto attaccato ha diversi problemi metodologici. In primo luogo, il segnale registrato è una misura indiretta di attività. Il segnale che viene misurato (come la cosiddetta azione in corso) è la corrente capacitiva, che spese la membrana (13). Si tratta di una questione estremamente importante metodologico. La capacità e la resistenza di una pipetta di registrazione in grado di filtrare il segnale registrato. E 'molto probabile che le correnti piccola azione si perdono nella carica la capacità della pipetta che, non possono essere adeguatamente compensata con maggior parte degli amplificatori, a causa della resistenza del headstage. Quando questi segnali non essere rilevati, il modello cottura apparente del neurone non riflette il vero modello di cottura. Allo stesso modo, non compensata pipetta e resistenze sigillo causare errori significativi nelle misurazioni durante i cambiamenti come quando le correnti di azione sono espressi (13). Alcuni amplificatori fornire capacità e della resistenza "compensazione" per la pipetta e la guarnizione, che limita la perdita di segnale, ma ad alta resistenza fasi capo della maggior parte degli amplificatori ostacolare compensazione ottimale. In secondo luogo, una situazione artificiale è imposto sulla cellula. In voltage-clamp modalità, l'area intorno alla membrana cellulare è tenuto a un potenziale fisso, in questi studi, 0 mV. Questo non significa che non c'è corrente applicata alla membrana cellulare. Il segnale misurato in voltage-clamp è in realtà la quantità di corrente applicata alla membrana per mantenere il potenziale fisso. Pertanto, questa corrente applicata può alterare l'attività delle cellule.
Registrazioni doppia nel sistema del GnRH sono particolarmente impegnativo a causa del numero limitato di neuroni GnRH e la loro distribuzione diffusa. Per le registrazioni doppio per avere successo, il manipolatore deve essere estremamente stabile. Anche leggero movimento degli elettrodi può causare la pipetta di scivolare fuori dal neurone e terminare la registrazione. Inoltre, il movimento della pipetta sulla cellula (per esempio, ri-posizionamento per compensare il movimento) possono alterare gli schemi di tiro. Alcuni canali ionici quali di tipo N canali del calcio sono meccanicamente sensibili: tratto membrana cause attività ripetitive in entrambe le configurazioni di registrazione intero-cellula e cellula-attached (14). Infine, il sistema manipolatore deve essere in grado di movimento estremamente sottile e liscia. Come notato sopra, con le registrazioni doppio, si prende due pipette alla superficie dei due neuroni selezionati in precedenza allo stesso tempo e cerca di sigillareun neurone. In caso di successo, poi si cerca di sigillare la seconda cella. In generale, non si può aspettare per sigillare e avere una registrazione di alta qualità con ogni tentativo. Questo, però, crea un problema particolare con le registrazioni doppio. Se uno è successo con il primo neurone ma non riesce con il secondo, si deve cambiare la pipetta e provare una cella diversa. Pertanto, si deve essere in grado di muoversi sia l'obiettivo ad immersione del microscopio e la pipetta alla parte superiore della perfusione bene (per cambiare la pipetta) senza interrompere il neurone con successo sigillato.
Il nostro sviluppo e l'uso del cell-attached allentato per le registrazioni approccio duale è un importante progresso tecnico per lo studio dei neuroni GnRH. E 'in grado di produrre risultati utili che aiuteranno spostare il campo in avanti nel contesto della questione critica di quali meccanismi alla base delle attività coordinata che si traduce nella secrezione ormonale pulsatile.
Acknowledgments
Sono grato a Ronald L. Calabrese, Dieter Jaeger (Emory University) e Ward Yuhas (Axon Instruments) per utili discussioni tecniche.
References
- Silverman, A. J. The gonadotropin releasing-hormone (GnRH) neuronal systems: immunocytochemistry. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Freeman, M. E. The neuroendocrine control of the ovarian cycle of the rat. In: The physiology of reproduction. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Belchetz, P. E., Plant, T. M., Nakai, Y., Keogh, E. J., Knobil, E. Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science. 202, 631-633 (1978).
- Maurer, J. A., Wray, S. Luteinizing hormone-releasing hormone quantified in tissues and slice explant cultures of postnatal rat hypothalami. Endocrinology. 140, 791-799 (1999).
- Harris, G. C., Levine, J. E. Pubertal acceleration of pulsatile gonadotropin-releasing hormone release in male rats as revealed by microdialysis. Endocrinology. 14, 163-171 (2003).
- Sisk, C. L., Richardson, H. N., Chappell, P. E., Levine, J. E. In vivo gonadotropin-releasing hormone secretion in female rats during peripubertal development and on proestrus. Endocrinology. 142, 2929-2936 (2001).
- Cardenas, H., Ordog, T., O'Byrne, K. T., Knobil, E. Single unit components of the hypothalamic multiunit electrical activity associated with the central signal generator that directs the pulsatile secretion of gonadotropic hormones. Proc Natl Acad Sci U S A. 90, 9630-9634 (1993).
- Silverman, A. J., Wilson, R., Kesner, J. S., Knobil, E. Hypothalamic localization of multiunit electrical activity associated with pulsatile LH release in the rhesus monkey. Neuroendocrinology. 44, 168-171 (1986).
- Roberts, C. B., O'Boyle, M. P., Suter, K. J. Dendrites determine the contribution of after depolarization potentials (ADPs) to generation of repetitive action potentials in hypothalamic gonadotropin releasing-hormone (GnRH) neurons. J Comput Neurosci. 26, 39-53 (2009).
- Spergel, D. J., Kruth, U., Hanley, D. F., Sprengel, R., Seeburg, P. H. GABA- and glutamate-activated channels in green fluorescent protein-tagged gonadotropin-releasing hormone neurons in transgenic mice. J Neurosci. 19, 2037-2050 (1999).
- Nelson, J. F., Felicio, L. S., Randall, P. K., Sims, C., Finch, C. E. A longitudinal study of estrous cyclicity in aging C57BL/6J mice: I. Cycle frequency, length and vaginal cytology. Biol Reprod. 27, 327-339 (1982).
- Gee, D. M., Flurkey, K., Finch, C. E. Aging and the regulation of luteinizing hormone in C57BL/6J mice: impaired elevations after ovariectomy and spontaneous elevations at advanced ages. Biol Reprod. 28, 598-607 (1983).
- Anson, B. D., Roberts, W. M. Loose-patch voltage-clamp technique. Neuromethods, Patch-clamp analysis: Advanced techniques. Walz, W., Boulton, A. A., Baker, G. B. 35, Humana Press. Place Unknown. 265-286 (2002).
- Calabrese, B., Tabarean, T., Juranka, P., Morris, C. E. Mechanosensitivity of N-type calcium channel currents. Biophys J. 83, 2560-2574 (2002).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Rapid killing of single neurons by irradiation of intracellularly injected dye. Science. 206, 702-704 (1979).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Lucifer Yellow slows voltage-gated Na+ current inactivation in a light-dependent manner in mice. J Physiol. 550, 159-167 (2003).

{kind=link}