

Summary
La actividad en los sistemas neuronales a menudo requiere descarga sincrónica del potencial de acción de las neuronas dentro de una población específica. Por ejemplo, los pulsos de hormona liberadora de gonadotropina (GnRH) es probable que requieran actividad coordinada entre las neuronas GnRH. Presentamos nuestro enfoque metodológico para la obtención simultánea de forma fiable los registros electrofisiológicos de las neuronas GnRH distribuye de manera difusa.
Abstract
Hormona liberadora de gonadotropina (GnRH) es un neuropéptido que regula la pequeña liberación pituitaria de la hormona luteinizante (LH) y hormona folículo-estimulante (FSH). Estas gonadotropinas son esenciales para la regulación de la función reproductiva. Las neuronas GnRH que contienen se distribuyen de forma difusa en todo el hipotálamo y el proyecto a la eminencia media, donde la liberación de GnRH desde sus terminales de los axones en el sistema portal hypophysiotropic (1). En los capilares portal, GnRH viaja a la glándula pituitaria anterior para estimular la liberación de las gonadotropinas en la circulación sistémica. La liberación de GnRH no es continua sino que ocurre en pulsos episódicos. Está bien establecido que la forma intermitente de la liberación de GnRH es esencial para la reproducción (2, 3).
Coordinación de la actividad de las neuronas GnRH probablemente múltiples pulsos de GnRH subyace. Contenido total de péptidos en las neuronas GnRH es de aproximadamente 1,0 pg / célula (4), de los cuales el 30% de probabilidades comprende la piscina liberable. Los niveles de GnRH durante un pulso (5, 6), sugieren múltiples neuronas de GnRH están probablemente involucradas en neurosecreción. Del mismo modo, la actividad de una sola unidad extraída de la unidad hipotálamo múltiples grabaciones durante la liberación de LH indica cambios en la actividad de múltiples neuronas (7). Los electrodos con la actividad registrada durante los pulsos de LH están asociados con somata GnRH o fibras (8). Por lo tanto, al menos parte de esta actividad surge de las neuronas GnRH.
Los mecanismos que resultan en el despido sincronizada de las neuronas GnRH del hipotálamo son desconocidos. Elucidar los mecanismos que coordinan el fuego en las neuronas GnRH es un problema complejo. En primer lugar, las neuronas GnRH son relativamente pocos en número. En los roedores, hay neuronas GnRH 800-2500. No está claro que todas las neuronas GnRH están involucrados en la liberación de GnRH episódica. Por otra parte, las neuronas GnRH se distribuye de manera difusa (1). Esto ha complicado nuestra comprensión de la coordinación de los disparos y ha hecho que muchos enfoques técnicos insuperables. Hemos optimizado suelta celular conectado a las grabaciones en el modo actual de mordaza para la detección directa de potenciales de acción y ha desarrollado un enfoque de grabación que permite la grabación simultánea de pares de neuronas GnRH.
Protocol
- Rebanadas hipotálamo del cerebro se preparan, se incuban y se transfiere a la cámara de grabación como se ha detallado anteriormente (9) en los animales, cuyas neuronas GnRH expresan la proteína verde fluorescente (GFP) bajo el control del promotor del péptido GnRH (10). Después de la extracción del cerebro del animal, el cerebro se bloquea con una cuchilla de afeitar para eliminar las zonas que no son de su interés. Con el fin de determinar las regiones a eliminar, el cerebro se coloca en la superficie dorsal de manera que el hipotálamo puede ser visualizada. Por tanto las orientaciones corte coronal y sagital, el cerebelo se ha eliminado. Para la preparación corte coronal, la porción rostral de la corteza se extrae. En esta orientación rebanada, la región lateral de la corteza proporcionan apoyo para cortar el cerebro y proporcionar suficiente área del cerebro para la colocación de alambres de plata para asegurar que el cerebro en la cámara de grabación. Para la preparación corte sagital, las regiones laterales del cerebro se eliminan y la parte rostral de la corteza proporcionan el apoyo para el cerebro de corte y la colocación de alambres de plata para asegurar que el cerebro en la grabación también. A lo largo del procedimiento de bloqueo, el cerebro está continuamente humedecido con el líquido frío artificial cefalorraquídeo (ACSF) utilizando una pipeta de vidrio.
- Una pequeña cantidad de pegamento se coloca en la plataforma de corte. El cerebro se levanta con una espátula. El exceso de ACSF se elimina por traer un Kimwipe a la superficie de la espátula y el dibujo ACSF sobrante del cerebro. El cerebro es suavemente se deslizó hacia delante en la cola. Es importante no tocar el cerebro o el empuje hacia abajo en él con el fin de asegurarla. El peso del cerebro en sí es suficiente para garantizar su seguridad.
- El cerebro y la plataforma de corte entonces se asegura que el microtomo vibración y el corte y se llena de frío ACSF. La solución fría de ambas empresas en el cerebro y puede proteger de posibles daños durante el procedimiento de corte. Diferentes regiones del cerebro varían en el grado de frialdad que optimiza la calidad de las rebanadas. La temperatura de la ACSF para la preparación de cortes hipotálamo es generalmente de 0 ° C.
- Lonchas deben ser cortadas y poco a poco cada corte debe flotar libremente fuera del cerebro. Si los segmentos empiezan a curvarse durante el corte, la velocidad a la que la hoja se debe avanzar más lento. Se puede utilizar un pincel pequeño para desenroscar rodajas, pero esto es el riesgo de dañar los cortes y por lo tanto, debe ser evitado. Poco a poco la velocidad de avance de la hoja es preferible utilizar un pincel.
- Como cada rodaja se corta en el cerebro, se retira de la cámara de corte y se coloca en una cámara de incubación de división en un baño de agua caliente (a unos 32 ° C). La porción de la incubadora es un suministro continuo de oxígeno, usando una mezcla de 95% O 2 y el 5% de CO 2. En general, el registro de pre-incubación oscila corte de 30 minutos a 2 horas.
- Pipetas de vidrio (3.6 MW) se fabrican como se describió previamente (9) con un extractor vertical. La temperatura y la duración de tirar varía en función del tipo de pipeta extractor se utiliza, el tipo de vidrio se utiliza y la vida útil del filamento de calentamiento del tirador. La forma de la punta de la pipeta también puede variar dependiendo de la preferencia del usuario. Hemos logrado buenos resultados con una punta afilada de manera uniforme con una abertura de 0,1 mm. Aberturas más grandes en la pipeta tienden a dañar las neuronas mientras que las pequeñas aberturas no culminan en grabaciones de larga duración.
- Pipetas recubiertos con Sylgard para reducir la capacidad. Sacó punta de la pipeta se ha cubierto ligeramente con Sylgard 184 con una pipeta segundos. Consejos Sylgarded son entonces curado con calor con una pistola de calor. Estas pipetas se llenan con la perfusión de baño. Para aumentar la visualización de la punta de la pipeta, una pequeña cantidad de Alexa-568 se añade a la parte de la solución del baño para ser usado como la solución de la pipeta antes de llenar las pipetas. No pesan los Alexa-568, sino más bien el uso suficiente para cambiar el color de rosa solución a simple vista. La punta de la pipeta puede ser visualizado en el filtro de Texas rojo.
- Hemos desarrollado un método para obtener corriente-clamp grabaciones para la detección de potenciales de acción con los sellos de baja resistencia (ver Discusión). Las grabaciones se realizaron en el potencial endógeno de reposo de la célula con Axoclamp 2B no aplica actual Instrumento con Axon. Sin embargo, debido al sello de baja resistencia, no se podía detectar los potenciales de acción con un solo amplificador 2B. Para aumentar la señal, un amplificador de segunda (AM Sistemas de 3000) se utiliza en la serie con el 2B Axoclamp. Este enfoque tiene la facilidad de sellos sueltos y la utilidad de registros a largo plazo. Técnicamente, se puede medir directamente los potenciales de acción, que permite eludir los problemas con el enfoque en el modo de grabación de voltaje-clamp (ver Discusión).
- Se toma dos pipetas de la superficie de las dos neuronas previamente seleccionados, al mismo tiempo, de una manera similar como el enfoque para la configuración de la celda grabación completa. Acercarse a los neurons al mismo tiempo, ha tenido más éxito en la obtención de las grabaciones de dos de tratar cada neurona de la pareja de forma independiente. Durante este tiempo, pulsos de corriente de inyección se pasan desde el amplificador 2B como se hace durante la aproximación para el conjunto de células grabaciones.
- La presión positiva se debe mantener en los dos pipetas para evitar la obstrucción de la punta. Presión positiva para las grabaciones de dos es el mejor generada mediante el uso de una jeringa de 3 ml sin llenar vacíos adjunta al titular de la pipeta por una longitud de la pieza de la tubería. Con jeringas permite mantener la presión en ambos pipetas. La presión positiva se debe aplicar tan pronto como la pipeta se ha colocado en la solución del baño de la cámara de grabación. Uno se acerca a las neuronas seleccionadas con las pipetas de forma secuencial. Después de colocar la primera pipeta directamente sobre la mayor superficie de la neurona, uno mantiene positiva, dejando la jeringa en su lugar y sale de la pipeta en esta posición. Uno vuelve a la parte superior de la cámara de registro y disminuye la pipeta segunda grabación en su posición. Neuronas seleccionadas están a menudo en los diferentes niveles de la división. La primera pipeta debe ser colocado sobre la neurona que es la más profunda en el corte y la posición de la pipeta segundo en la neurona más superficial. Esto evita que el posicionamiento de la pipeta segundo de cambiar el posicionamiento de la pipeta segundo que la división se moverá ligeramente en el plano vertical como pipetas entrar en el sector.
- Sellos sueltos (18-30 MW) se utilizará. La presión positiva se induce un pequeño hoyuelo en la superficie de cada célula. Uno los intentos de cerrar la primera neurona por la liberación de presión positiva y la aplicación de presión negativa muy leve. La presión positiva se libera mediante la eliminación de la jeringa. De succión con la boca se aplica al extremo libre de la tubería. La membrana de una neurona sana responderá rápidamente a la liberación de presión positiva y se adhieran a la pipeta. Ligera succión se forma un sello con la resistencia adecuada en una neurona sana.
- Cuando uno intenta sellar una neurona y no, la pipeta no se puede volver a utilizar ya que las membranas sólo sello muy limpio de vidrio. En cambio, eleva la pipeta (y objetivo de microscopio) de distancia de la superficie de la porción de la parte superior de la perfusión, así, elimina la pipeta del baño, los cambios en una pipeta limpia y luego regresa a la corte para tratar de sellar otra célula . Por estas razones, es importante seleccionar múltiples neuronas potencial para las grabaciones. Para cortes con menos de 4.6 neuronas calidad de grabación entre los que elegir, el éxito en la obtención de grabaciones de doble es limitado.
- Después de establecer un sello suelto sobre la base de la resistencia, los pulsos de inyección de corriente del amplificador 2B se terminan. El amplificador 2B se coloca en serie con el amplificador de la serie 3000. En esta configuración, la salida del amplificador 2B sirve como entrada para el amplificador de la serie 3000. El amplificador proporciona una señal de éste para la adquisición de datos.
- El sello (paso 4) y la reconfiguración de amplifers (paso 5) se repite para la segunda neurona.

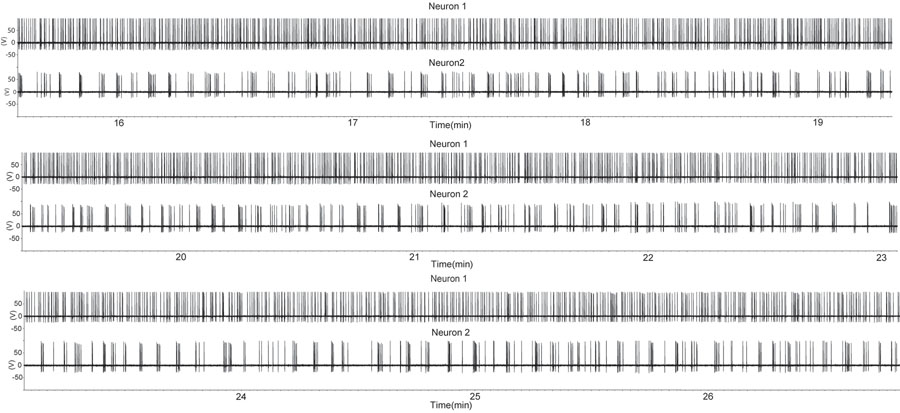
Figura 1. Un período de aumento de disparar en dos neuronas GnRH con la configuración de grabación del sello suelta adjunta en una preparación de corte sagital. El corte se obtuvo de un macho castrado. Cada flecha hacia arriba indica un potencial de acción. Tenga en cuenta que una neurona GnRH (huellas parte superior) muestra disparando casi continua, mientras que la segunda neurona GnRH presenta ráfagas intermitentes. Este patrón de actividad de las neuronas GnRH solo es similar a la de las unidades individuales extraídos de grabaciones de varias unidades en la secreción de hormonas en vivo (7). Por favor, haga clic aquí para ver una versión ampliada de la figura 1.
Discussion
La actividad de interés en algunas neuronas incluidas las neuronas GnRH (basado en la secreción de hormonas) se produce en escalas de tiempo de hora (5-7). Por lo tanto, la configuración de célula entera no es la mejor opción para algunos de los objetivos experimentales, debido a la diálisis de mensajeros intracelulares en el modo de grabación de toda la célula. En segundo lugar, de células enteras grabaciones se limitan generalmente a animales de menos de 120 días de edad. Con la edad, las membranas neuronales parecen rígidos, por lo que los sellos de alta resistencia difícil de lograr. Además, si se obtiene un sello de alta resistencia, rompiendo el parche rompe el sello, dejando un espacio entre la pipeta y la membrana. Esto conduce a una grabación quede inutilizable y una neurona que morirá rápidamente debido a los desequilibrios iónicos. Ciclos regulares de ovario, y por lo tanto, la actividad estable del generador de pulsos de GnRH no se produce hasta más tarde en la vida (7-10 meses de edad en las mujeres C57BL6, 11, 12), más allá de la edad en que uno puede razonablemente anticipar la obtención de células enteras grabaciones de forma fiable. Finalmente, de células enteras grabaciones destruir las relaciones endógenas de las concentraciones de iones internos y externos. Con las grabaciones de células enteras, la concentración interna de cualquier ion es igual a la concentración de los iones en la solución de la pipeta. Esto es así porque el volumen de la solución de la pipeta es relativamente grande alcanza rápidamente el equilibrio con / reemplaza el volumen endógeno relativamente pequeño de la célula.
El enfoque de células vinculada suelta evita muchas de las limitaciones de células enteras grabaciones. En primer lugar, un sello de baja resistencia (15-30 MW) se puede utilizar. Estos son relativamente fáciles de formar, incluso en las neuronas de los animales más viejos. En segundo lugar, uno no se rompe el parche de membrana sellada. Por lo tanto, las grabaciones sueltas de células vinculada son técnicamente mucho más fácil que su conjunto de células grabaciones. Además, desde la membrana celular está intacta, la diálisis de los componentes intracelulares no ocurre y endógenos relaciones iónicas se conservan. No se puede utilizar el enfoque de células vinculada sueltas para el estudio de las corrientes sinápticas, pero es ideal para los registros a largo plazo de las neuronas de una manera relativamente no invasiva. Las grabaciones de las células adjunto también se puede realizar utilizando cualquier solución intracelular estándar en la pipeta. Esto ofrece la ventaja adicional de la ruptura de la membrana de parche cuando la grabación se completa a largo plazo y el etiquetado de la neurona con un marcador intracelular.
El enfoque de células vinculada sueltas se ha utilizado en el modo de grabación de voltaje-clamp. Sin embargo, el voltaje-clamp grabación en la configuración de la celda adjunta suelta tiene varios problemas metodológicos. En primer lugar, la señal grabada es una medida indirecta de la actividad. La señal que se mide (como la acción de llamada actual) es la corriente capacitiva que los cargos de la membrana (13). Se trata de una cuestión metodológica muy importante. La capacidad y la resistencia de una pipeta de grabación se puede filtrar la señal grabada. Es muy probable que las corrientes de acción pequeños se pierden en la carga de la capacidad de la pipeta, que no pueden ser compensados con la mayoría de los amplificadores, debido a la alta resistencia de la headstage. Cuando estas señales no se detectan, el patrón de activación aparente de la neurona no se corresponde con el patrón de activación real. Del mismo modo, la pipeta no compensada y resistencias sello causar errores significativos en las mediciones durante los cambios, como cuando las corrientes de acción se expresan (13). Algunos amplificadores proporcionan la capacidad y resistencia a la "compensación" por la pipeta y el sello, lo que limita la pérdida de señal, pero de alta resistencia a la cabeza de las etapas de la mayoría de los amplificadores de obstaculizar la compensación óptima. En segundo lugar, una situación artificial, se impone a la célula. En el modo de voltaje-clamp, el área alrededor de la membrana celular se mantiene a un potencial fijo, en estos estudios, 0 mV. Esto no significa que no hay corriente aplicada a la membrana celular. La señal de voltaje-clamp es en realidad la cantidad de corriente aplicada a la membrana para mantener el potencial fijo. Por lo tanto, esta corriente aplicada puede alterar la actividad de las células.
Grabaciones de doble en el sistema de GnRH son particularmente difíciles debido al número limitado de neuronas GnRH y su distribución difusa. Para las grabaciones de doble para tener éxito, el manipulador debe ser muy estable. Incluso un ligero movimiento del electrodo puede causar la pipeta para deslizarse fuera de la neurona y finalizar la grabación. Por otra parte, el movimiento de la pipeta de la célula (por ejemplo, el cambio de posición para compensar el movimiento) puede alterar los patrones de activación. Algunos canales iónicos como el tipo N-canales de calcio son mecánicamente sensibles: estirar la membrana hace que la actividad repetitiva en las configuraciones de grabación tanto de células enteras y de células vinculada (14). Por último, el sistema de manipulación debe ser capaz de movimiento excesivamente fina y suave. Como se señaló anteriormente, con las grabaciones de dos, se toma dos pipetas de la superficie de las dos neuronas previamente seleccionados, al mismo tiempo y los intentos de cerraruna neurona. Si tiene éxito, entonces los intentos de cerrar la segunda celda. Por lo general, no se puede esperar para sellar y tienen una alta calidad de grabación con cada intento. Sin embargo, esto crea un problema particular con las grabaciones de doble. Si uno tiene éxito con la primera neurona, pero no con el segundo, se debe cambiar la pipeta y tratar de una celda diferente. Por lo tanto, uno debe ser capaz de mover tanto el objetivo de inmersión del microscopio y de la pipeta a la parte superior de la perfusión y (para cambiar la pipeta) sin interrumpir la neurona con éxito sellado.
Nuestro desarrollo y uso de la suelta de células adjunta enfoque dual de las grabaciones es un gran avance técnico en el estudio de las neuronas GnRH. Es probable que produzca resultados útiles que le ayudarán a avanzar este campo en el contexto de la cuestión crítica de qué mecanismos subyacen a la actividad coordinada que se traduce en la secreción hormonal pulsátil.
Acknowledgments
Doy las gracias a Ronald L. Calabrese, Dieter Jaeger (Emory University) y Ward Yuhas (Axon Instruments) para útiles discusiones técnicas.
References
- Silverman, A. J. The gonadotropin releasing-hormone (GnRH) neuronal systems: immunocytochemistry. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Freeman, M. E. The neuroendocrine control of the ovarian cycle of the rat. In: The physiology of reproduction. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Belchetz, P. E., Plant, T. M., Nakai, Y., Keogh, E. J., Knobil, E. Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science. 202, 631-633 (1978).
- Maurer, J. A., Wray, S. Luteinizing hormone-releasing hormone quantified in tissues and slice explant cultures of postnatal rat hypothalami. Endocrinology. 140, 791-799 (1999).
- Harris, G. C., Levine, J. E. Pubertal acceleration of pulsatile gonadotropin-releasing hormone release in male rats as revealed by microdialysis. Endocrinology. 14, 163-171 (2003).
- Sisk, C. L., Richardson, H. N., Chappell, P. E., Levine, J. E. In vivo gonadotropin-releasing hormone secretion in female rats during peripubertal development and on proestrus. Endocrinology. 142, 2929-2936 (2001).
- Cardenas, H., Ordog, T., O'Byrne, K. T., Knobil, E. Single unit components of the hypothalamic multiunit electrical activity associated with the central signal generator that directs the pulsatile secretion of gonadotropic hormones. Proc Natl Acad Sci U S A. 90, 9630-9634 (1993).
- Silverman, A. J., Wilson, R., Kesner, J. S., Knobil, E. Hypothalamic localization of multiunit electrical activity associated with pulsatile LH release in the rhesus monkey. Neuroendocrinology. 44, 168-171 (1986).
- Roberts, C. B., O'Boyle, M. P., Suter, K. J. Dendrites determine the contribution of after depolarization potentials (ADPs) to generation of repetitive action potentials in hypothalamic gonadotropin releasing-hormone (GnRH) neurons. J Comput Neurosci. 26, 39-53 (2009).
- Spergel, D. J., Kruth, U., Hanley, D. F., Sprengel, R., Seeburg, P. H. GABA- and glutamate-activated channels in green fluorescent protein-tagged gonadotropin-releasing hormone neurons in transgenic mice. J Neurosci. 19, 2037-2050 (1999).
- Nelson, J. F., Felicio, L. S., Randall, P. K., Sims, C., Finch, C. E. A longitudinal study of estrous cyclicity in aging C57BL/6J mice: I. Cycle frequency, length and vaginal cytology. Biol Reprod. 27, 327-339 (1982).
- Gee, D. M., Flurkey, K., Finch, C. E. Aging and the regulation of luteinizing hormone in C57BL/6J mice: impaired elevations after ovariectomy and spontaneous elevations at advanced ages. Biol Reprod. 28, 598-607 (1983).
- Anson, B. D., Roberts, W. M. Loose-patch voltage-clamp technique. Neuromethods, Patch-clamp analysis: Advanced techniques. Walz, W., Boulton, A. A., Baker, G. B. 35, Humana Press. Place Unknown. 265-286 (2002).
- Calabrese, B., Tabarean, T., Juranka, P., Morris, C. E. Mechanosensitivity of N-type calcium channel currents. Biophys J. 83, 2560-2574 (2002).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Rapid killing of single neurons by irradiation of intracellularly injected dye. Science. 206, 702-704 (1979).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Lucifer Yellow slows voltage-gated Na+ current inactivation in a light-dependent manner in mice. J Physiol. 550, 159-167 (2003).

{kind=link}