

Summary
L'activité dans les systèmes neuronaux nécessite souvent des décharges action synchrone potentiels de neurones dans une population spécifique. Par exemple, des impulsions de la gonadolibérine (GnRH) nécessitera probablement une activité coordonnée entre les neurones à GnRH. Nous présentons notre approche méthodologique pour l'obtention simultanée de manière fiable des enregistrements électrophysiologiques des neurones GnRH diffusément réparti.
Abstract
Gonadolibérine (GnRH) est un neuropeptide qui régule la libération petite glande pituitaire de l'hormone lutéinisante (LH) et hormone folliculo-stimulante (FSH). Ces gonadotrophines sont essentielles pour la régulation de la fonction de reproduction. Les neurones contenant le GnRH sont distribuées de façon diffuse à travers l'hypothalamus et de projets à l'éminence médiane où ils libèrent la GnRH de leurs terminaisons axonales dans le système porte hypophysiotropic (1). Dans les capillaires du portail, la GnRH se rend à l'hypophyse antérieure pour stimuler la libération des gonadotrophines dans la circulation systémique. GnRH n'est pas continue mais se produit plutôt en impulsions épisodiques. Il est bien établi que la manière intermittente de la libération de GnRH est essentiel pour la reproduction (2, 3).
La coordination de l'activité des neurones à GnRH multiples sous-tend probablement impulsions GnRH. Peptide contenu dans les neurones à GnRH totale est d'environ 1,0 pg / cellule (4), dont 30% de chances comprend la piscine amovible. Niveaux de la GnRH pendant une impulsion (5, 6), suggèrent plusieurs neurones à GnRH sont probablement impliqués dans neurosécrétion. De même, l'activité seule unité extraite de l'hypothalamus à logements multiples enregistrements pendant la libération de LH indique les changements dans l'activité des neurones multiples (7). Les électrodes à l'activité enregistrée au cours des impulsions de LH sont associés soit à soma GnRH ou de fibres (8). Par conséquent, au moins certains de cette activité résulte de neurones à GnRH.
Les mécanismes qui aboutissent à feu synchronisées dans les neurones à GnRH hypothalamique sont inconnus. Élucider les mécanismes qui coordonnent tir dans les neurones à GnRH est un problème complexe. Tout d'abord, les neurones GnRH sont relativement peu nombreux. Chez les rongeurs, il ya des neurones à GnRH 800-2500. Il n'est pas clair que tous les neurones à GnRH sont impliqués dans la libération de GnRH épisodique. Par ailleurs, les neurones à GnRH sont distribuées de façon diffuse (1). Cela a compliqué notre compréhension de la coordination des tirs et a fait de nombreuses approches techniques insolubles. Nous avons optimisé perdre cellule attachée enregistrements en courant en mode pince pour la détection directe des potentiels d'action et a développé une approche qui permet l'enregistrement des enregistrements simultanés de paires de neurones à GnRH.
Protocol
- Tranches de cerveau hypothalamique sont préparés, incubé et transféré à la chambre d'enregistrement comme précédemment détaillées (9) chez les animaux dont les neurones expriment la GnRH protéine fluorescente verte (GFP) sous le contrôle du promoteur du peptide GnRH (10). Après l'élimination du cerveau de l'animal, le cerveau est bloquée à l'aide d'une lame de rasoir pour éliminer les régions qui ne sont pas d'intérêt. Afin de déterminer les régions de supprimer, le cerveau est placé sur la surface dorsale de sorte que l'hypothalamus peut être visualisée. Pour les deux orientations tranche coronale et sagittale, le cervelet est enlevé. Pour préparation de tranches coronales, la partie rostrale du cortex est enlevé. Dans cette orientation tranche, les régions latérales du cortex fournir un soutien pour le tranchage du cerveau et de fournir suffisamment d'espace de cerveau pour le placement des fils d'argent pour sécuriser le cerveau dans la chambre d'enregistrement. Pour la préparation coupe sagittale, les régions latérales du cerveau sont enlevés et les portions rostrales du cortex fournir le soutien pour le cerveau de tranchage et de placement des fils d'argent pour sécuriser le cerveau à l'enregistrement bien. Tout au long de la procédure de blocage, le cerveau est constamment humidifié par le froid liquide céphalorachidien artificiel (ACSF) à l'aide d'une pipette en verre.
- Une petite quantité de superglue est placé sur la plate-forme de coupe. Le cerveau est soulevé avec une spatule. L'excès est éliminé par l'ACSF apporter une Kimwipe à la surface de la spatule et le dessin ACSF excès loin du cerveau. Le cerveau est doucement glissé vers l'avant sur la colle. Il est important de ne pas toucher le cerveau ou le pousser vers le bas sur elle afin de la sécuriser. Le poids du cerveau lui-même est généralement suffisant pour s'assurer qu'il est sécuritaire.
- Le cerveau et la plate-forme de coupe est alors fixé à la microtome vibrant et la coupe est bien remplie avec le froid ACSF. La solution à froid à la fois le cerveau et les entreprises peuvent protéger contre les dommages causés lors de la procédure de découpe. Différentes régions du cerveau varient selon le degré de froideur qui optimise la qualité des tranches. La température de l'ACSF pour la préparation des tranches hypothalamiques est généralement de 0 ° C.
- Tranches doit être coupé lentement et chaque tranche devrait flotter librement loin du cerveau. Si les tranches commencent à se courber pendant la coupe, la vitesse à laquelle la lame est la promotion devrait être ralenti. On peut utiliser un petit pinceau à des tranches de déplier, mais cela ne risque d'endommager les tranches et donc, devrait être évitée. Lentement, la vitesse d'avance de la lame est préférée à l'aide d'un pinceau.
- Comme chaque tranche est coupée par le cerveau, il est retiré de la chambre de coupe et placé dans une chambre d'incubation tranche dans un bain d'eau tiède (environ 32 ° C). L'incubateur tranche est alimenté en continu avec de l'oxygène en utilisant un mélange de 95% d'O 2 et 5% de CO 2. Généralement, le pré-enregistrement d'incubation varie tranche de 30 minutes à 2 heures.
- Pipettes en verre (3-6 MQ) sont fabriquées comme décrit précédemment (9) en utilisant un extracteur vertical. La température et la durée de tirant varie en fonction du type de pipette extracteur on utilise, le type de verre, on utilise et de la durée de vie du filament de chauffage de l'extracteur. La forme de la pointe de la pipette peut aussi varier selon les préférences de l'utilisateur. Nous avons obtenu de bons résultats avec une pointe effilée de manière uniforme avec une ouverture de 0,1 mm. Des ouvertures plus grandes dans la pipette ont tendance à endommager les neurones alors que les petites ouvertures n'aboutissent pas à des enregistrements de longue durée.
- Pipettes enduits de Sylgard de réduire la capacité. Embouts de pipette sont tiré légèrement enduit d'Sylgard 184 secondes avec une pipette. Conseils Sylgarded sont ensuite durcis en utilisant la chaleur d'un pistolet thermique. Ces pipettes sont ensuite remplies avec le perfusat bain. Pour augmenter la visualisation des embouts de pipette, une petite quantité d'Alexa-568 est ajoutée à la portion de la solution de bain pour être utilisé comme la solution la pipette avant de remplir les pipettes. Nous ne pèsent pas l'Alexa-568, mais plutôt l'utilisation suffit de tourner le rose solution en vue. Les embouts de pipette peuvent ensuite être visualisées sous le filtre rouge Texas.
- Nous avons développé une approche pour obtenir des enregistrements de courant pince pour la détection des potentiels d'action en utilisant des joints à faible résistance (voir discussion). Les enregistrements sont effectués à endogènes de la cellule potentiel de repos avec Axoclamp 2B pas appliquée actuel Instrument Axon aide de. Toutefois, en raison de l'étanchéité à faible résistance, on ne pouvait pas détecter les potentiels d'action avec juste l'amplificateur 2B. Pour augmenter le signal, un second amplificateur (AM Systems 3000) est utilisé en série avec le 2B Axoclamp. Cette approche a de la facilité de joints lâches et l'utilité des enregistrements à long terme. Techniquement, on peut mesurer directement les potentiels d'action, contournant ainsi les problèmes liés à l'approche dans le mode d'enregistrement voltage-clamp (voir discussion).
- On prend deux pipettes à la surface des deux neurones préalablement sélectionné dans le même temps, d'une façon similaire à l'approche de la configuration de l'enregistrement de cellules entières. En approchant les deux neurons dans le même temps a été plus réussi à obtenir les enregistrements doubles que de tenter chaque neurone de la paire de manière indépendante. Pendant ce temps, les impulsions d'injection de courant sont transmis de l'amplificateur 2B comme cela est fait lors de l'approche des cellules entières d'enregistrements.
- Une pression positive doit être maintenue sur les deux pipettes pour éviter le colmatage de la pointe. Une pression positive pour les enregistrements doubles est le meilleur produit en utilisant une seringue de 3 ml vacants vide attaché à la porte-pipette par une longueur de morceau de tuyau. Utiliser des seringues permet de maintenir la pression sur les deux pipettes. Une pression positive doit être appliqué dès que la pipette a été placé dans la solution du bain de la chambre d'enregistrement. Une des approches les neurones sélectionnés avec les pipettes de manière séquentielle. Après le positionnement de la première pipette directement sur la plus grande surface du neurone, on maintient positifs en laissant la seringue en place et laisse la pipette dans cette position. On retourne alors à la partie supérieure de la chambre d'enregistrement et abaisse la pipette deuxième enregistrement en position. Neurones sont souvent choisis à différents niveaux de la tranche. La première pipette devrait être positionné sur le neurone qui est le plus profond dans la tranche et la position de la pipette deuxième neurone plus superficielles. Cela empêche le positionnement de la pipette seconde de se déplacer le positionnement de la pipette seconde que la tranche se déplace légèrement dans le plan vertical que les pipettes entrer dans la tranche.
- Joints lâches (18-30 MQ) sera utilisé. La pression positive va induire une petite fossette sur la surface de chaque cellule. Une tentative de sceller le premier neurone en libérant la pression positive et l'application très légère pression négative. La pression positive est libéré en retirant la seringue. Aspiration par la bouche est alors appliquée à l'extrémité libre de la tubulure. Un neurone membrane saine sera de répondre rapidement à la libération de pression positive et adhèrent à la pipette. Légère aspiration va former un joint avec une résistance appropriée dans un neurone sain.
- Lorsque l'on tente de sceller un neurone et échoue, la pipette ne peut pas être ré-utilisé depuis des membranes d'étanchéité à seulement très propre verre. Au lieu de cela, on élève la pipette (et un objectif de microscope) loin de la surface de la tranche au sommet de la perfusion ainsi, élimine la pipette du bain, des modifications à une pipette propre, puis revient à la tranche d'essayer de sceller une autre cellule . Pour ces raisons, il est important de sélectionner plusieurs neurones potentiels pour les enregistrements. Pour les tranches de 4-6 avec moins de neurones qualité de l'enregistrement à partir de laquelle choisir, la réussite dans l'obtention des enregistrements doubles est limité.
- Après l'établissement d'un joint lâche basée sur la résistance, les impulsions d'injection de courant issu de l'amplificateur 2B sont terminées. L'amplificateur 2B est placée en série avec l'amplificateur de la série 3000. Dans cette configuration, la sortie de l'amplificateur 2B sert d'entrée pour l'amplificateur de la série 3000. L'amplificateur de signal pour ce dernier fournit d'acquisition de données.
- Le sceau (étape 4) et la reconfiguration de amplifers (étape 5) est ensuite répété pour la deuxième neurone.

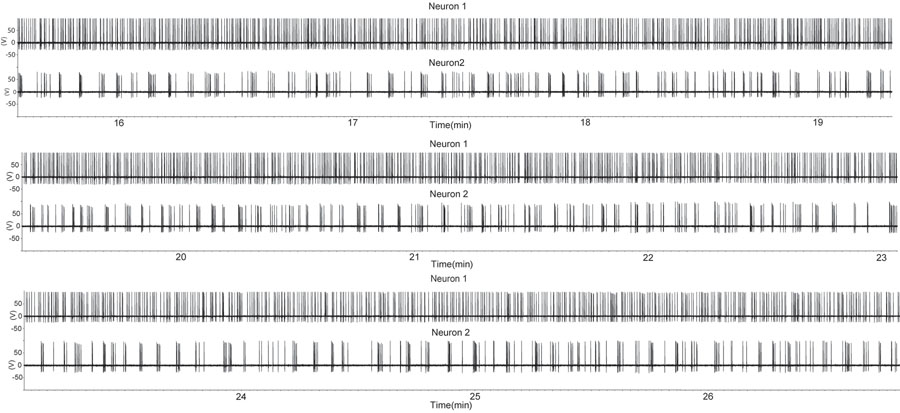
Figure 1. Une période de mise à feu a augmenté dans deux neurones à GnRH en utilisant la configuration d'enregistrement joint lâches attachées dans une préparation coupe sagittale. La tranche a été dérivé à partir d'un mâle castré. Chaque flèche vers le haut indique un potentiel d'action. Notez qu'un neurone GnRH (traces en haut) montre tir presque continu tandis que le second neurone GnRH expositions rafales intermittentes. Ce modèle d'activité de neurones à GnRH seule est similaire à celui des unités simples extraits de plusieurs unités d'enregistrements pendant la sécrétion d'hormone in vivo (7). S'il vous plaît cliquez ici pour voir une version agrandie de la figure 1.
Discussion
L'activité d'intérêt dans certains neurones dont les neurones à GnRH (basé sur la sécrétion d'hormones) se produit sur des échelles de temps d'heures (5-7). Par conséquent, la configuration cellule entière n'est pas le meilleur choix pour certains objectifs expérimentaux en raison de la dialyse de messagers intracellulaires dans le mode d'enregistrement de cellules entières. Secondairement, la cellule entière enregistrements sont généralement limités aux animaux de moins de 120 jours d'âge. Avec l'âge, les membranes neuronales semblent durcir, rendant joints à haute résistance difficile à réaliser. De plus, si on obtient un joint à haute résistance, la rupture du patch perturbe le sceau, laissant un trou entre la pipette et la membrane. Cela conduit à un enregistrement inutilisable et un neurone qui meurent rapidement à cause de déséquilibres ioniques. Cycles ovariens régulière, et donc, une activité stable du générateur d'impulsions GnRH n'a pas lieu avant tard dans la vie (7-10 mois d'âge dans C57BL6 femelles; 11, 12), au-delà de l'âge où l'on peut raisonnablement prévoir l'obtention de cellules entières d'enregistrements de manière fiable. Enfin, la cellule entière enregistrements détruire les ratios des concentrations en ions endogènes internes et externes. Avec les enregistrements de cellules entières, la concentration interne de tout ion est égale à la concentration de l'ion dans la solution pipette. C'est parce que le volume de la solution de pipette relativement important atteint rapidement l'équilibre avec / remplace le volume relativement faible endogène de la cellule.
L'approche cellulaire lâche attachée contourne plusieurs des limites de la cellule entière enregistrements. Tout d'abord, un joint à faible résistance (15-30 MQ) peuvent être utilisés. Ils sont relativement faciles à former, même dans les neurones provenant d'animaux âgés. Deuxièmement, on ne rompt pas le patch de membrane étanche. Par conséquent, les enregistrements de cellules lâches attachées sont techniquement beaucoup plus facile que la cellule entière enregistrements. En outre, depuis la membrane cellulaire est intacte, la dialyse de composants intracellulaires ne se produit pas et endogène des ratios ioniques sont préservés. On ne peut pas utiliser l'approche cellulaire lâche-joint pour étudier les courants synaptiques mais il est idéal à long terme des enregistrements de neurones d'une manière relativement non invasive. Les enregistrements cellule attachée peut également être effectuée en utilisant une solution standard de intracellulaire dans la pipette. Ceci offre l'avantage supplémentaire de la rupture du patch de membrane lorsque l'enregistrement à long terme est terminée et l'étiquetage du neurone avec un marqueur intracellulaire.
L'approche cellulaire lâche-joint a été utilisé dans le mode d'enregistrement voltage-clamp. Toutefois, voltage-clamp enregistrement dans la configuration de la cellule lâche-joint a plusieurs problèmes méthodologiques. Tout d'abord, le signal enregistré est une mesure indirecte de l'activité. Le signal qui est mesuré (comme l'action dite de courant) est le courant capacitif que les frais de la membrane (13). C'est une question extrêmement importante méthodologiques. La capacité et la résistance d'une pipette d'enregistrement peut filtrer le signal enregistré. Il est très probable que les courants d'action de petites sommes perdus dans la capacité de charge de la pipette qui ne peuvent pas être correctement indemnisés pour la plupart des amplificateurs, en raison de la forte résistance de l'headstage. Lorsque ces signaux ne sont pas détectés, la tendance apparente de feu le neurone ne reflète pas la tendance tirs vrai. De même, une pipette non compensée et résistances joint entraîner des erreurs significatives dans les mesures lors des changements tels que lorsque les courants d'action sont exprimés (13). Certains amplificateurs fournissent la capacité et la résistance «compensation» pour la pipette et le sceau, ce qui limite la perte de signal, mais la tête haute résistance des stades de la plupart des amplificateurs d'entraver une compensation optimale. Deuxièmement, une situation artificielle est imposée sur la cellule. En voltage-clamp mode, la zone autour de la membrane cellulaire est tenu à un potentiel fixe, dans ces études, 0 mV. Cela ne signifie pas l'absence de courant appliqué à la membrane cellulaire. Le signal mesuré en voltage-clamp est en fait la quantité de courant appliqué à la membrane pour maintenir le potentiel fixe. Par conséquent, ce courant appliqué peut altérer l'activité cellulaire.
Enregistrements dans le système de double GnRH sont particulièrement difficiles en raison du nombre limité de neurones à GnRH et de leur distribution diffuse. Pour les enregistrements doubles pour réussir, le manipulateur doit être extrêmement stable. Même un léger mouvement de l'électrode peut causer la pipette à glisser le neurone et la fin de l'enregistrement. Par ailleurs, le mouvement de la pipette sur la cellule (par exemple, re-positionnement pour compenser le mouvement) peut modifier les modèles de tir. Certains canaux ioniques tels les canaux calciques de type N sont mécaniquement sensibles: étirer la membrane provoque activité répétitive dans des configurations d'enregistrement à la fois toute-cellule et cellule-joint (14). Enfin, le système manipulateur doit être capable de mouvement excessivement fine et lisse. Comme indiqué plus haut, avec des enregistrements doubles, on prend deux pipettes à la surface des deux neurones préalablement sélectionné dans le même temps et les tentatives pour scellerun neurone. En cas de succès, alors on tente de sceller la deuxième cellule. Généralement, on ne peut pas s'attendre à avoir un sceau et d'enregistrement de haute qualité à chaque tentative. Ceci, cependant, pose un problème particulier avec les enregistrements doubles. Si l'on est réussi avec le premier neurone, mais échoue à la seconde, il faut changer la pipette et d'essayer une autre cellule. Par conséquent, on doit être capable de se déplacer à la fois l'objectif à immersion du microscope et de la pipette vers le haut de la perfusion ainsi (pour changer la pipette) sans perturber le neurone succès scellés.
Notre développement et l'utilisation des lâches cellule attachée approche pour les enregistrements double est une avancée technique majeure dans l'étude de neurones à GnRH. Il est susceptible de produire des résultats utiles qui aideront le champ en avant dans le contexte de la question critique de ce qui sous-tendent les mécanismes de l'activité coordonnée qui résulte de la sécrétion de l'hormone pulsatile.
Acknowledgments
Je suis reconnaissant à Ronald L. Calabrese, Dieter Jaeger (Emory University) et Ward Yuhas (Axon Instruments) pour des discussions techniques utiles.
References
- Silverman, A. J. The gonadotropin releasing-hormone (GnRH) neuronal systems: immunocytochemistry. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Freeman, M. E. The neuroendocrine control of the ovarian cycle of the rat. In: The physiology of reproduction. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Belchetz, P. E., Plant, T. M., Nakai, Y., Keogh, E. J., Knobil, E. Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science. 202, 631-633 (1978).
- Maurer, J. A., Wray, S. Luteinizing hormone-releasing hormone quantified in tissues and slice explant cultures of postnatal rat hypothalami. Endocrinology. 140, 791-799 (1999).
- Harris, G. C., Levine, J. E. Pubertal acceleration of pulsatile gonadotropin-releasing hormone release in male rats as revealed by microdialysis. Endocrinology. 14, 163-171 (2003).
- Sisk, C. L., Richardson, H. N., Chappell, P. E., Levine, J. E. In vivo gonadotropin-releasing hormone secretion in female rats during peripubertal development and on proestrus. Endocrinology. 142, 2929-2936 (2001).
- Cardenas, H., Ordog, T., O'Byrne, K. T., Knobil, E. Single unit components of the hypothalamic multiunit electrical activity associated with the central signal generator that directs the pulsatile secretion of gonadotropic hormones. Proc Natl Acad Sci U S A. 90, 9630-9634 (1993).
- Silverman, A. J., Wilson, R., Kesner, J. S., Knobil, E. Hypothalamic localization of multiunit electrical activity associated with pulsatile LH release in the rhesus monkey. Neuroendocrinology. 44, 168-171 (1986).
- Roberts, C. B., O'Boyle, M. P., Suter, K. J. Dendrites determine the contribution of after depolarization potentials (ADPs) to generation of repetitive action potentials in hypothalamic gonadotropin releasing-hormone (GnRH) neurons. J Comput Neurosci. 26, 39-53 (2009).
- Spergel, D. J., Kruth, U., Hanley, D. F., Sprengel, R., Seeburg, P. H. GABA- and glutamate-activated channels in green fluorescent protein-tagged gonadotropin-releasing hormone neurons in transgenic mice. J Neurosci. 19, 2037-2050 (1999).
- Nelson, J. F., Felicio, L. S., Randall, P. K., Sims, C., Finch, C. E. A longitudinal study of estrous cyclicity in aging C57BL/6J mice: I. Cycle frequency, length and vaginal cytology. Biol Reprod. 27, 327-339 (1982).
- Gee, D. M., Flurkey, K., Finch, C. E. Aging and the regulation of luteinizing hormone in C57BL/6J mice: impaired elevations after ovariectomy and spontaneous elevations at advanced ages. Biol Reprod. 28, 598-607 (1983).
- Anson, B. D., Roberts, W. M. Loose-patch voltage-clamp technique. Neuromethods, Patch-clamp analysis: Advanced techniques. Walz, W., Boulton, A. A., Baker, G. B. 35, Humana Press. Place Unknown. 265-286 (2002).
- Calabrese, B., Tabarean, T., Juranka, P., Morris, C. E. Mechanosensitivity of N-type calcium channel currents. Biophys J. 83, 2560-2574 (2002).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Rapid killing of single neurons by irradiation of intracellularly injected dye. Science. 206, 702-704 (1979).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Lucifer Yellow slows voltage-gated Na+ current inactivation in a light-dependent manner in mice. J Physiol. 550, 159-167 (2003).

{kind=link}