

Summary
Деятельность в нейронных систем часто требует синхронной потенциала действия разрядов от нейронов в конкретной группе населения. Например, бобовые гонадотропин-рилизинг гормона (ГнРГ), вероятно, потребует скоординированных действий между ГнРГ нейронов. Мы представляем наш методологический подход к получению надежного одновременного электрофизиологических записей с диффузно распределенных ГнРГ нейронов.
Abstract
Гонадотропин-рилизинг гормона (ГнРГ) представляет собой небольшой нейропептида, который регулирует гипофиз релизе лютеинизирующего гормона (ЛГ) и фолликулостимулирующего гормона (ФСГ). Эти гонадотропинов имеют важное значение для регуляции репродуктивной функции. GnRH-содержащие нейроны распределяются диффузно по всему гипоталамус и проект срединного возвышения, где они выпускают ГнРГ от своих терминалов аксона в hypophysiotropic портальной системе (1). В портала капилляров, GnRH едет в передней доли гипофиза, чтобы стимулировать высвобождение гонадотропинов в системный кровоток. GnRH релиз не является непрерывным, а происходит в эпизодических импульсов. Хорошо известно, что прерывистый способ ГнРГ релиз имеет важное значение для воспроизводства (2, 3).
Координация деятельности нескольких нейронов ГнРГ, вероятно, лежит в основе импульсов ГнРГ. Общее содержание пептида в ГнРГ нейронов составляет приблизительно 1,0 пг / клетка (4), из которых 30% вероятно, включает в себя разборных бассейн. Уровни ГнРГ во время импульса (5, 6), предлагаю несколько ГнРГ нейронов, вероятно, участвует в нейросекреции. Кроме того, одна единица активности, извлеченные из гипоталамуса многоквартирных записи во время LH релизе указывает изменения в деятельности нескольких нейронов (7). Электроды с записанными активность во время импульсов ЛГ связаны либо с ГнРГ somata или волокон (8). Поэтому, по крайней мере часть этой деятельности возникает из-ГнРГ нейронов.
Механизмы, которые приводят к синхронизированной стрельбы в нейронах гипоталамуса ГнРГ неизвестны. Выяснению механизмов, которые координируют стрельбы в ГнРГ нейронов представляет собой сложную проблему. Во-первых, ГнРГ нейронов относительно мало. У грызунов, Есть 800-2500 ГнРГ нейронов. Пока не ясно, что все ГнРГ нейронов участвуют в эпизодических релиз ГнРГ. Более того, ГнРГ нейронов диффузно распределенные (1). Это усложнило наше понимание координации стрельбы и сделал много технических подходов неразрешимыми. Мы оптимизировали свободные ячейки подключением записей в текущем режиме зажим для прямого выявления потенциалов действия и развитых записи подход, который позволяет одновременно записи из пар ГнРГ нейронов.
Protocol
- Гипоталамус мозга ломтиками готовы, инкубируют и переданы записи камеры, как ранее подробным (9) в животных, ГнРГ нейронов выразить зеленый флуоресцентный белок (GFP) под контролем промотора ГнРГ пептид (10). После удаления мозга от животных, мозг заблокированы с помощью лезвия бритвы для устранения регионов, которые не представляют интереса. Для того чтобы определить регионы, чтобы удалить, мозг размещении на ее спинной поверхности, так что гипоталамус может быть визуализированы. Для обеих корональные и сагиттального ориентации ломтик, мозжечок удален. Для подготовки корональных ломтик, ростральной части коры снимается. В этом срез ориентации, боковых отделах коры оказывать поддержку для нарезки мозга и обеспечить достаточную область мозга для размещения серебряные провода для обеспечения мозга в записи камеры. Для подготовки сагиттальном срезе, боковых участках мозга удаляются и ростральной части коры обеспечивают поддержку мозга нарезки и для размещения проводов серебра для обеспечения мозга в записи хорошо. На протяжении блокирование процедуры, мозг постоянно смоченную холодной искусственной цереброспинальной жидкости (ACSF) с использованием стеклянной пипетки.
- Небольшое количество суперклей находится на переднем платформы. Мозг поднимается с помощью шпателя. Превышение ACSF удаляется принести Kimwipe на поверхность шпателем и рисования избыток ACSF от мозга. Мозг мягко скользила вперед на клей. Важно, чтобы не трогать мозг или надавите на нее для того, чтобы обеспечить это. Вес самого мозга, как правило, достаточно, чтобы убедиться, что он в безопасности.
- Мозга и резки платформы затем фиксируется в вибрирующих микротома и резки яма заполняется холодной ACSF. Холодный раствор обеих фирм мозга и может защитить от повреждений во время резки процедуры. Различные области мозга различаются по степени холодности, которая оптимизирует качество ломтиками. Температура ACSF для подготовки гипоталамуса ломтиками, как правило, 0 ° C.
- Ломтики должны быть сокращены медленно и каждый кусочек должно свободно плавать вдали от мозга. Если ломтики начинают скручиваться во время резки, скорость, с которой лезвие продвижения должны быть замедлены. Можно использовать маленькую кисть, чтобы развиваться ломтиками но это риск повреждения ломтиками и, таким образом, следует избегать. Постепенно скорость продвижения лезвия является предпочтительным по сравнению с использованием кисти.
- Поскольку каждый срез вырезать из мозга, он снимается с режущей камеры и помещают в камеру ломтик инкубации в теплую ванну водой (примерно 32 ° С). Ломтик инкубаторе постоянно снабжаются кислородом с использованием смеси 95% O 2 и 5% СО 2. Как правило, предварительная запись ломтик инкубации составляет от 30 минут до 2 часов.
- Стекло пипетки (3-6 МОм) изготавливаются, как описано выше (9), используя вертикальные съемника. Температуры и длительности потянув изменяется в зависимости от типа пипетки съемник используются, типа стекла один использует и жизни нагрева нити съемника. Форму кончика пипетки также может варьироваться в зависимости от предпочтений пользователя. Мы добились хороших успехов равномерно конический наконечник с отверстием 0,1 мм. Большие отверстия в пипетку, как правило, повреждение нейронов, в то время как меньшие отверстия не в состоянии привести к длительным записи продолжительностью.
- Пипетки покрытые Sylgard уменьшить емкость. Тянули наконечники для пипеток слегка покрытые Sylgard 184, используя второй пипетки. Sylgarded советы, то тепловой сушки при помощи тепловой пушки. Эти пипетки затем заполняют ванну перфузат. Для увеличения визуализации наконечники для пипеток, небольшое количество Alexa-568 добавляется в часть ванны решение для использования в качестве пипетки раствор до заполнения пипеток. Мы не весят Alexa-568, а использовать достаточно, чтобы превратить решение розовые на глаз. Наконечники могут быть визуализированы в Техасе красный фильтр.
- Мы разработали подход для получения текущей записи-зажим для обнаружения потенциалов действия с использованием низких печатей сопротивления (см. Обсуждение). Записи выполняются на эндогенный потенциал покоя клетка с 2B Axoclamp не применяется тока с помощью Axon инструмента. Однако из-за низкого сопротивления печать, никто не мог обнаружить потенциалов действия с помощью всего 2В усилителя. Для увеличения сигнала, второго усилителя (АМ Системы 3000) используется в серии с 2B Axoclamp. Этот подход имеет легкость свободной печати и полезность долгосрочного записей. Технически, можно непосредственно измерить потенциалы действия, тем самым обойти проблемы с подходом в напряжении зажим режиме записи (см. Обсуждение).
- Один берет два пипетки на поверхность двух ранее выбранных нейронов в то же время, таким же образом, как подход для всей конфигурации записи клетки. Подойдя как пеurons в то же время были более успешными в получении двойной записи не пытаться каждый нейрон пары самостоятельно. В течение этого времени импульсов тока инъекции передаются от 2В усилителя, как это делается во время подхода к цельноклеточная записей.
- Положительное давление должно поддерживаться на обоих пипетки для предотвращения засорения наконечника. Положительное давление на двойной записи лучше формируется с использованием незаполненных 3 мл пустой шприц прилагается к пипетки держатель длина кусок трубы. Использование шприцев позволяет сохранять давление на обе пипеток. Положительное давление должно применяться, как только пипеткой был помещен в ванну решение записи камеры. Один подходы выбранных нейронов с пипетки последовательно. После установки первой пипетки прямо над крупнейшим поверхности нейрона, один поддерживает положительный, оставляя шприц на месте и листьев пипеткой в этом положении. Один затем возвращается в начало записи камеры и снижает второй пипетки записи в нужное положение. Выбранный нейроны часто на разных уровнях среза. Первый пипетки должны быть расположены на нейрон, что является самым глубоким в срезе и положение второй пипетки на более поверхностном нейрона. Это предотвращает позиционирование второй пипетки от сдвига позиционирование второй пипетки, как часть будет немного двигаться в вертикальной плоскости, что пипетки ввести срез.
- Свободные печатей (18-30 МОм) будет использоваться. Положительное давление будет вызывать небольшие лунки на поверхности каждой клетки. Один попытки печать первого нейрона, выпустив положительного давления и применения очень небольшое отрицательное давление. Положительное давление будет снято, удалив шприц. Всасывающая ртом Затем на свободный конец трубки. Мембраны здоровых нейронов будет оперативно реагировать на выпуск положительное давление и придерживаться пипетки. Незначительное всасывание сформирует печать с соответствующим сопротивлением в здоровых нейронов.
- Когда одна попытка, чтобы запечатать нейрона и не удается, пипетки не могут быть повторно использованы после мембраны только уплотнение очень чистый стакан. Вместо этого повышает пипетки (и объектива микроскопа) от поверхности среза в начало перфузии хорошо, снимает пипетки из ванны, изменения в чистой пипеткой, а затем возвращается в часть, чтобы попытка изолировать другую ячейку . По этим причинам, важно, чтобы выбрать несколько потенциальных нейронов для записей. Для срезов менее 4-6 качество записи нейронов из которых можно выбирать, успех в получении двойных записей ограничено.
- После установления свободную печать основана на сопротивление, импульсы тока инжекции с 2В усилителя прекращается. Amplifer 2B помещается в серии с серии 3000 amplifer. В этой конфигурации, выход amplifer 2B служит входом для серии 3000 amplifer. Последний amplifer обеспечивает сигнал для сбора данных.
- Уплотнение (шаг 4) и реконфигурации amplifers (шаг 5) повторяется для второго нейрона.

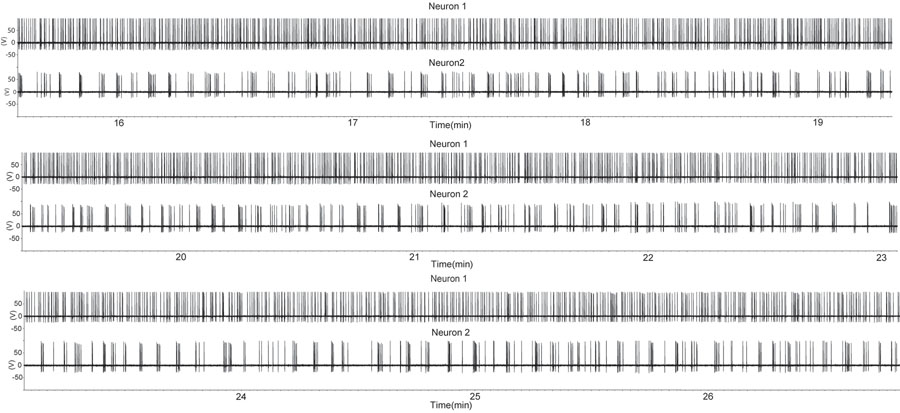
Рисунок 1. Период увеличилась стрельбы в двух ГнРГ нейронов использованием свободно печать прилагается конфигурации записи в сагиттальной подготовки срез. Часть была получена из кастрированный. Каждое отклонение вверх указывает потенциала действия. Отметим, что один ГнРГ нейронов (вверху следы) экспонатов почти непрерывной стрельбы в то время как второй ГнРГ нейронов экспонатов прерывистый всплесков. Такой характер деятельности от одного ГнРГ нейронов же, как отдельных единиц, извлеченные из многоквартирных записи во время секрецию гормона в естественных условиях (7). Пожалуйста, нажмите здесь , чтобы видеть большую версию рисунке 1.
Discussion
Деятельности, представляющей интерес в некоторых нейронов в том числе ГнРГ нейронов (основано на секрецию гормонов) происходит на временных масштабах часов (5-7). Таким образом, вся конфигурация ячейки не лучший выбор для некоторых экспериментальных целей из-за диализ внутриклеточных мессенджеров в целом режим записи клетки. Во вторую очередь, цельноклеточная записи, как правило, ограничивается животных меньше, чем о 120-дневного возраста. С возрастом появляются нейрональных мембран напрягаться, делая высокой печати сопротивления трудно достичь. Кроме того, если один получает высокую печать сопротивление, разрывая патч нарушает печать, оставляя отверстие между пипеткой и мембраны. Это приводит к непригодным для использования записи и нейрона, который позволяет быстро умирают из-за ионного дисбаланса. Регулярные циклы яичников, и, таким образом, стабильная деятельность ГнРГ генератора импульсов происходит только в более позднем возрасте (7-10 месяцев в C57Bl6 самок, 11, 12), вне возраста, когда можно обоснованно ожидать получения цельноклеточная записей надежно. Наконец, цельноклеточная записей уничтожить эндогенных соотношения внутренних и внешних концентраций ионов. В целом записи клетки, внутренней концентрации любого иона равна концентрации ионов в пипетку раствор. Это потому, что решение пипетки сравнительно большой объем быстро достигает равновесия с / заменяет относительно небольшой объем эндогенные клетки.
Свободные ячейки прилагается подход позволяет обойти многие ограничения цельноклеточная записей. Во-первых, низкая печать сопротивления (15-30 МОм) могут быть использованы. Они относительно легко даже в форме нейронов с более старых животных. Во-вторых, никто не разрыв запечатанных участок мембраны. Таким образом, свободная клетка прилагается записей технически гораздо проще, чем цельноклеточная записей. Кроме того, так как клеточная мембрана не повреждена, диализ внутриклеточных компонентов не происходит, и эндогенные ионных отношения сохраняются. Нельзя использовать свободные ячейки прилагается подход к изучению синаптические токи, но он идеально подходит для долгосрочной записи из нейронов в относительно неинвазивным способом. Клеточного прилагается записи могут также быть выполнены с использованием любого стандартного внутриклеточного раствора в пипетку. Это дает дополнительное преимущество разрывом мембраны патч, когда продолжительной записи срок завершения и маркировки нейрон с внутриклеточных маркер.
Свободные ячейки прилагается подход был использован в напряжении зажим режиме записи. Тем не менее, напряжение-зажим записи в свободные ячейки прилагается конфигурация имеет ряд методологических проблем. Во-первых, записанный сигнал косвенным показателем активности. Сигнал, который измеряется (как так называемые действия тока) емкостной ток, который заряжает мембрану (13). Это чрезвычайно важный методологический вопрос. Емкость и сопротивление записи пипетки может фильтровать записанного сигнала. Очень вероятно, что малые токи действия теряются в зарядке емкость пипетки, которая, не может быть должным образом компенсирован у большинства усилителей, из-за высокой устойчивости headstage. Когда эти сигналы остаются незамеченными, очевидно шаблон стрельбы нейрона не отражает истинной картины стрельбы. Кроме того, некомпенсированные пипетки и печатью сопротивления привести к значительным ошибкам в измерениях во время изменения, такие, как тогда, когда токи действия выражаются (13). Некоторые усилители обеспечивают емкость и сопротивление "компенсации" за пипетку и печатью, что ограничивает потери сигнала, но и высокой устойчивостью голову стадиях большинство усилителей препятствует оптимальной компенсации. Во-вторых, искусственная ситуация накладывается на ячейку. В напряжения зажим режим, область вокруг клеточных мембран проводится для фиксированного потенциала, в этих исследованиях, 0 мВ. Это не означает, в настоящее время нет применительно к клеточной мембране. Сигнал измеряется в вольт-зажим на самом деле количество тока применяются к мембраны для поддержания фиксированного потенциала. Таким образом, это касалось ток может изменять активность клеток.
Двойной записи в ГнРГ системы особенно сложно из-за ограниченного числа ГнРГ нейронов и их диффузное распределение. Для двойной записи, чтобы быть успешным, манипулятор должна быть чрезвычайно стабильна. Даже незначительные движения электрода может вызвать пипетку соскользнуть нейрона и окончания записи. Более того, движение пипетки на ячейку (например, повторное позиционирование для компенсации движения) может изменить стрельбы узоры. Некоторые ионные каналы, такие N-типа кальциевых каналов механически чувствительных: мембранные растянуть причины повторяющихся деятельности как целых клеток и клеточных прилагаются записи конфигурации (14). Наконец, манипулятор система должна быть способна чрезвычайно тонкой и плавного движения. Как отмечалось выше, с двумя записями, одна принимает два пипетки на поверхность двух ранее выбранных нейронов в то же время и пытается печатьодин нейрон. В случае успеха, то попытки, чтобы запечатать второй ячейке. Вообще, нельзя ожидать, чтобы запечатать и имеют высокое качество записи с каждой попыткой. Это, однако, создает особую проблему с двойной записи. Если кто-то успешно с первого нейрона, но не со второго, надо менять пипетки и попробуйте другой ячейке. Таким образом, надо уметь передвигаться как погружение цель микроскопа и пипеткой в начало перфузии также (чтобы изменить пипетки), не нарушая успешно запечатанных нейрона.
Наши разработки и использования свободной ячейки подключением подход к двойной записи является значительным техническим достижением в изучении ГнРГ нейронов. Это, вероятно, принести полезные результаты, которые помогут двигаться вперед поле в контексте критических вопрос о том, какие механизмы лежат в основе скоординированной деятельности, что приводит к пульсирующей секреции гормона.
Acknowledgments
Я благодарен Рональд Л. Калабрезе, Дитер Jaeger (Университет Эмори) и Уорд Юхас (Axon Instruments) за полезные обсуждения технических вопросов.
References
- Silverman, A. J. The gonadotropin releasing-hormone (GnRH) neuronal systems: immunocytochemistry. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Freeman, M. E. The neuroendocrine control of the ovarian cycle of the rat. In: The physiology of reproduction. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Belchetz, P. E., Plant, T. M., Nakai, Y., Keogh, E. J., Knobil, E. Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science. 202, 631-633 (1978).
- Maurer, J. A., Wray, S. Luteinizing hormone-releasing hormone quantified in tissues and slice explant cultures of postnatal rat hypothalami. Endocrinology. 140, 791-799 (1999).
- Harris, G. C., Levine, J. E. Pubertal acceleration of pulsatile gonadotropin-releasing hormone release in male rats as revealed by microdialysis. Endocrinology. 14, 163-171 (2003).
- Sisk, C. L., Richardson, H. N., Chappell, P. E., Levine, J. E. In vivo gonadotropin-releasing hormone secretion in female rats during peripubertal development and on proestrus. Endocrinology. 142, 2929-2936 (2001).
- Cardenas, H., Ordog, T., O'Byrne, K. T., Knobil, E. Single unit components of the hypothalamic multiunit electrical activity associated with the central signal generator that directs the pulsatile secretion of gonadotropic hormones. Proc Natl Acad Sci U S A. 90, 9630-9634 (1993).
- Silverman, A. J., Wilson, R., Kesner, J. S., Knobil, E. Hypothalamic localization of multiunit electrical activity associated with pulsatile LH release in the rhesus monkey. Neuroendocrinology. 44, 168-171 (1986).
- Roberts, C. B., O'Boyle, M. P., Suter, K. J. Dendrites determine the contribution of after depolarization potentials (ADPs) to generation of repetitive action potentials in hypothalamic gonadotropin releasing-hormone (GnRH) neurons. J Comput Neurosci. 26, 39-53 (2009).
- Spergel, D. J., Kruth, U., Hanley, D. F., Sprengel, R., Seeburg, P. H. GABA- and glutamate-activated channels in green fluorescent protein-tagged gonadotropin-releasing hormone neurons in transgenic mice. J Neurosci. 19, 2037-2050 (1999).
- Nelson, J. F., Felicio, L. S., Randall, P. K., Sims, C., Finch, C. E. A longitudinal study of estrous cyclicity in aging C57BL/6J mice: I. Cycle frequency, length and vaginal cytology. Biol Reprod. 27, 327-339 (1982).
- Gee, D. M., Flurkey, K., Finch, C. E. Aging and the regulation of luteinizing hormone in C57BL/6J mice: impaired elevations after ovariectomy and spontaneous elevations at advanced ages. Biol Reprod. 28, 598-607 (1983).
- Anson, B. D., Roberts, W. M. Loose-patch voltage-clamp technique. Neuromethods, Patch-clamp analysis: Advanced techniques. Walz, W., Boulton, A. A., Baker, G. B. 35, Humana Press. Place Unknown. 265-286 (2002).
- Calabrese, B., Tabarean, T., Juranka, P., Morris, C. E. Mechanosensitivity of N-type calcium channel currents. Biophys J. 83, 2560-2574 (2002).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Rapid killing of single neurons by irradiation of intracellularly injected dye. Science. 206, 702-704 (1979).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Lucifer Yellow slows voltage-gated Na+ current inactivation in a light-dependent manner in mice. J Physiol. 550, 159-167 (2003).

{kind=link}