

Summary
Atividade neuronal em sistemas muitas vezes exige descargas ação síncrona potencial de neurônios dentro de uma população específica. Por exemplo, pulsos de hormônio liberador de gonadotrofinas (GnRH) provavelmente exigirá atividade coordenada entre os neurônios GnRH. Apresentamos nossa abordagem metodológica para a obtenção simultânea confiável gravações eletrofisiológicas dos neurônios GnRH difusamente distribuídas.
Abstract
Hormônio Liberador de Gonadotropina (GnRH) é um neuropeptídeo de pequeno porte que regula a liberação hipofisária do hormônio luteinizante (LH) e hormônio folículo estimulante (FSH). Estes gonadotrofinas são essenciais para a regulação da função reprodutiva. Os neurônios GnRH contendo estão distribuídos difusamente pelo hipotálamo e projeto para a eminência mediana onde liberam GnRH a partir de seus terminais de axônios no sistema portal hypophysiotropic (1). Nos capilares portal, GnRH viaja para a glândula hipófise para estimular a liberação de gonadotrofinas na circulação sistêmica. Liberação de GnRH não é contínua, mas ocorre em pulsos episódica. Está bem estabelecido que a maneira intermitente de liberação de GnRH é essencial para a reprodução (2, 3).
Coordenação da atividade dos neurônios GnRH, provavelmente, vários pulsos de GnRH está subjacente. Peptídeo conteúdo total em neurônios GnRH é de aproximadamente 1,0 pg / célula (4), dos quais 30% de probabilidade compreende o conjunto liberável. Níveis de GnRH durante um pulso (5, 6), sugerem múltiplos neurônios GnRH provavelmente estão envolvidos na Neurossecreção. Da mesma forma, a atividade única unidade extraído hipotálamo multi-unit gravações durante a libertação de LH indica mudanças na atividade dos neurônios múltiplos (7). Os eletrodos com atividade registrada durante pulsos de LH estão associados com qualquer somata GnRH ou fibras (8). Portanto, pelo menos algumas dessas atividades decorre de neurônios GnRH.
Os mecanismos que resultam em demissão sincronizado nos neurônios GnRH hipotalâmico são desconhecidos. Elucidar os mecanismos que coordenam a disparar em neurônios GnRH é um problema complexo. Primeiro, os neurônios GnRH são relativamente poucos em número. Em roedores, há 800-2500 neurônios GnRH. Não é claro que todos os neurônios GnRH estão envolvidos na liberação de GnRH episódica. Além disso, os neurônios GnRH são difusamente distribuídos (1). Isso tem complicado a nossa compreensão da coordenação de queima e tem feito muitas abordagens técnicas intratável. Nós otimizamos solta células anexado gravações em pinça de corrente para o modo de detecção direta de potenciais de ação e desenvolveu uma abordagem de gravação que permite gravações simultâneas de pares de neurônios GnRH.
Protocol
- Fatias de cérebro do hipotálamo são preparadas, incubadas e transferidos para a câmara de gravação como anteriormente detalhada (9) em animais cujos neurônios GnRH expressam a proteína verde fluorescente (GFP) sob o controle do promotor peptídeo GnRH (10). Após a remoção do cérebro do animal, o cérebro é bloqueado usando uma lâmina de barbear para eliminar as regiões que não são de interesse. A fim de determinar as regiões de remover, o cérebro é colocado em sua superfície dorsal, para que o hipotálamo pode ser visualizado. Tanto para as orientações corte coronal e sagital, o cerebelo é removido. Para a preparação de corte coronal, a porção rostral do córtex é removido. Nesta orientação fatia, as regiões laterais do córtex fornecer suporte para cortar o cérebro e proporcionar área suficiente do cérebro para a colocação de fios de prata para proteger o cérebro na câmara de gravação. Para a preparação fatia sagital, as regiões laterais do cérebro são removidas e as porções rostral do córtex fornecer o apoio para o cérebro corte e para a colocação de fios de prata para proteger o cérebro na gravação também. Durante todo o processo de bloqueio, o cérebro está continuamente umedecida com o frio do líquido cefalorraquidiano artificial (ACSF) utilizando uma pipeta de vidro.
- Uma pequena quantidade de Superglue é colocado na plataforma de corte. O cérebro é levantado com uma espátula. ACSF excesso é removido por trazer um Kimwipe à superfície da espátula e desenho ACSF excesso longe do cérebro. O cérebro é suavemente deslizou em frente para a cola. É importante não tocar no cérebro ou empurrar para baixo sobre ele, a fim de prendê-lo. O peso do cérebro em si é geralmente suficiente para garantir que é seguro.
- Do cérebro e da plataforma de corte é então presa ao micrótomo vibrando e assim o corte é preenchido com ACSF frio. A solução fria ambas as empresas o cérebro e pode proteger contra danos durante o procedimento de corte. Diferentes regiões do cérebro variam no grau de frieza que otimiza fatias de qualidade. A temperatura do ACSF para a preparação de fatias de hipotálamo é geralmente 0 ° C.
- Fatias devem ser cortadas de forma lenta e cada fatia deve flutuar livremente longe do cérebro. Se começar a enrolar as fatias durante o corte, a velocidade com que a lâmina está avançando deve ser retardado. Pode-se usar um pincel pequeno para fatias uncurl mas este corre o risco de danificar as fatias e, portanto, deve ser evitado. Lentamente, a velocidade de avanço da lâmina é preferido sobre o uso de um pincel.
- Como cada fatia é cortada a partir do cérebro, é retirado da câmara de corte e colocados em uma câmara de incubação fatia em um banho de água morna (aproximadamente 32 ° C). A fatia incubadora é continuamente fornecido com oxigênio usando uma mistura de 95% O 2 e 5% CO 2. Geralmente, os pré-gravação varia fatia de incubação de 30 minutos a 2 horas.
- Pipetas de vidro (3-6 mohms) são fabricados conforme descrito anteriormente (9) com um puxador vertical. A temperatura ea duração de puxar varia de acordo com o tipo de pipeta utiliza um extrator, o tipo de vidro se usa eo tempo de vida do filamento de aquecimento do extrator. A forma da ponta da pipeta também pode variar, dependendo da preferência do usuário. Temos conseguido um bom sucesso com uma ponta uniformemente cônico com uma abertura de 0,1 mm. Aberturas maiores na pipeta tendem a danificar os neurônios enquanto aberturas menores não resultarem em gravações de longa duração.
- Pipetas revestido com Sylgard para reduzir a capacitância. Ponteiras são levemente puxados revestido com Sylgard 184 utilizando uma pipeta segundo. Dicas Sylgarded são, então, calor curado usando uma pistola de calor. Estas pipetas são preenchidos com o perfusato banho. Para aumentar a visualização das ponteiras, uma pequena quantidade de Alexa-568 é adicionado à parte da solução de banho para ser usado como a solução da pipeta antes de preencher as pipetas. Nós não pesar o Alexa-568, mas sim usar o suficiente para transformar o pink solução por olho. As pontas da pipeta pode ser visualizado sob o vermelho Texas filtro.
- Nós desenvolvemos uma abordagem para obter grampo corrente gravações para a detecção de potenciais de ação usando selos de baixa resistência (ver Discussão). As gravações são realizadas em potencial endógeno das células de descanso com Axoclamp 2B não aplicada Instrumento Axon atual usando o. No entanto, devido ao selo de baixa resistência, não se poderia detectar potenciais de ação com apenas o amplificador 2B. Para aumentar o sinal, um amplificador de segunda (AM Sistemas de 3000) é usado em série com a 2B Axoclamp. Esta abordagem tem a facilidade de selos soltos e utilidade de gravações a longo prazo. Tecnicamente, pode-se medir diretamente a potenciais de ação, contornando assim os problemas com a abordagem no modo de gravação de tensão-clamp (ver discussão).
- Um leva duas pipetas para a superfície dos dois neurônios previamente selecionados, ao mesmo tempo, de forma semelhante como a abordagem para a configuração de toda célula de gravação. Aproximando-se tanto neurons ao mesmo tempo, tem sido mais bem sucedido em obter as gravações dupla de tentar cada neurônio do par de forma independente. Durante este tempo, os pulsos de injeção de corrente são passadas do amplificador 2B como é feito durante a aproximação para a célula inteira gravações.
- Pressão positiva deve ser mantida em ambas as pipetas para evitar o entupimento da ponta. Pressão positiva para as gravações é a melhor dupla gerados pela utilização de uma seringa de 3 ml por preencher vazios anexados ao titular da pipeta por um período de pedaço de tubo. Uso de seringas permite manter a pressão sobre ambos os pipetas. Pressão positiva deve ser aplicada assim que a pipeta foi colocada na solução de banho da câmara de gravação. Se aborda os neurônios selecionados com as pipetas seqüencialmente. Depois de posicionar a pipeta primeiro diretamente sobre a maior superfície do neurônio, se mantém positivo, deixando a seringa no local e deixa a pipeta nesta posição. Um retorna ao topo da câmara de gravação e reduz a pipeta segunda gravação para a posição. Neurônios selecionados são, muitas vezes em diferentes níveis da fatia. A pipeta primeiro deve ser posicionado sobre o neurônio que é o mais profundo na fatia e posição da pipeta no segundo neurônio mais superficial. Isso evita que o posicionamento da pipeta segundo de mudar o posicionamento da pipeta segundo como a fatia irá mover-se ligeiramente no plano vertical como pipetas entrar na fatia.
- Selos soltos (18-30 mohms) será usado. A pressão positiva irá induzir uma pequena covinha na superfície de cada célula. Uma tentativa de selar o primeiro neurônio, liberando pressão positiva e aplicação muito ligeira pressão negativa. A pressão positiva é liberado por retirar a seringa. De sucção com a boca é então aplicado à extremidade livre do tubo. Um neurônio membrana saudável irá responder rapidamente à liberação de pressão positiva e aderir à pipeta. Ligeira sucção irá formar uma vedação com resistência adequada em um neurônio saudável.
- Quando se tenta selar um neurônio e falhar, a pipeta não pode ser reutilizado desde membranas só selo para muito limpa vidro. Em vez disso, uma levanta a pipeta (e objetiva do microscópio) de distância da superfície da fatia para o topo da perfusão bem, remove a pipeta do banho, muda para uma pipeta limpa e depois volta para a fatia para tentar selar outra célula . Por essas razões, é importante para selecionar vários neurônios potencial para as gravações. Para fatias com menos de 4-6 neurônios qualidade de gravação de que para escolher, o sucesso na obtenção de gravações dupla é limitada.
- Depois de estabelecer um selo soltos com base na resistência, os pulsos de injeção de corrente do amplificador 2B são encerrados. O amplifer 2B é colocado em série com o amplificador da série 3000. Nesta configuração, a saída do amplificador 2B serve como entrada para o amplificador da série 3000. O amplifer último fornece sinal para aquisição de dados.
- O selo (passo 4) e reconfiguração do amplificador de (passo 5) é então repetido para o segundo neurônio.

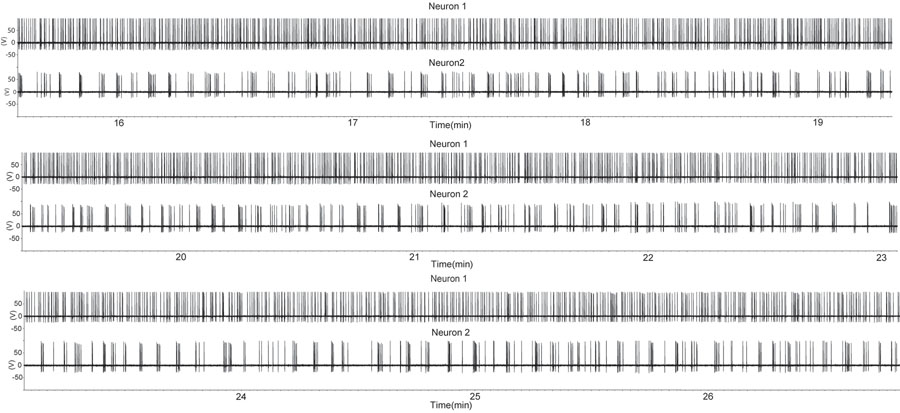
Figura 1. Um período de aumento de disparo em dois neurônios GnRH utilizando a configuração solta selo de gravação anexada em uma preparação fatia sagital. A fatia foi derivado de um macho castrado. Cada deflexão para cima indica um potencial de ação. Note-se que um neurônio GnRH (traços em cima) apresenta disparar quase contínua, enquanto o segundo GnRH neurônio apresenta rajadas intermitentes. Este padrão de atividade de neurônios individuais GnRH é semelhante ao de unidades individuais extraídas da unidade de multi-gravações durante a secreção hormonal in vivo (7). Por favor, clique aqui para ver uma versão ampliada da figura 1.
Discussion
A atividade de interesse em alguns neurônios, incluindo os neurônios GnRH (com base na secreção de hormônios) ocorre em escalas de tempo de horas (5-7). Portanto, a configuração da célula de todo não é a melhor escolha para algumas metas experimental devido à diálise de mensageiros intracelulares no modo de toda célula de gravação. Secundariamente, de célula inteira gravações são geralmente limitada a animais com menos de cerca de 120 dias de idade. Com a idade, membranas neuronais parecem endurecer, tornando vedações de alta resistência difícil de alcançar. Além disso, se obtém-se um selo de alta resistência, rompendo o patch rompe o selo, deixando um buraco entre a pipeta ea membrana. Isto leva a uma gravação inutilizável e um neurônio que rapidamente morrem devido a desequilíbrios iônicos. Ciclos ovariano regular, e, assim, a atividade estável do gerador de pulsos de GnRH não ocorre até mais tarde na vida (7-10 meses de idade em C57BL6 fêmeas, 11, 12), para além da idade quando se pode razoavelmente prever a obtenção de célula inteira gravações de forma confiável. Finalmente, de célula inteira gravações destruir as relações endógenas das concentrações dos íons internos e externos. Com gravações de células inteiras, a concentração interna de qualquer íon é igual à concentração do íon na solução da pipeta. Isso ocorre porque a solução de pipeta de volume relativamente grande rapidamente atinge o equilíbrio com / substitui o volume relativamente pequeno endógena da célula.
A abordagem de células soltas anexado contorna muitas das limitações de toda célula gravações. Primeiro, um selo de baixa resistência (15-30 mohms) pode ser usado. Estas são relativamente fáceis de forma ainda nos neurônios de animais mais velhos. Em segundo lugar, não é o patch ruptura da membrana selada. Portanto, as gravações de células soltas anexados são tecnicamente muito mais fácil do que de célula inteira gravações. Além disso, uma vez que a membrana celular está intacta, a diálise de componentes intracelulares não ocorre e endógenos relações iônicas são preservadas. Não se pode usar a abordagem de células soltas em anexo para estudar as correntes sinápticas mas é ideal para gravações a longo prazo dos neurônios de uma forma relativamente não-invasivo. As gravações de células-anexado também podem ser realizadas usando qualquer solução padrão intracelular na pipeta. Isto oferece a vantagem adicional de ruptura do remendo de membrana quando a gravação de longa duração está concluído e rotulagem do neurônio com um marcador intracelular.
A abordagem de células soltas anexado tem sido usada no modo de gravação de tensão-clamp. No entanto, a tensão de clamp de gravação na configuração da célula solta anexado tem vários problemas metodológicos. Primeiro, o sinal gravado é uma medida indireta de atividade. O sinal que é medido (como a ação chamada atual) é a corrente capacitiva que as taxas de membrana (13). Esta é uma questão extremamente importante metodológicas. A capacitância ea resistência de uma pipeta de gravação pode filtrar o sinal gravado. É muito provável que as correntes pequena ação são perdidos no carregamento da capacitância da pipeta que, não pode ser devidamente compensados com a maioria dos amplificadores, devido à alta resistência do headstage. Quando estes sinais passam despercebidos, o padrão de ativação aparente do neurônio não reflete o padrão de disparo verdade. Do mesmo modo, pipetar descompensada e resistências selo causar erros significativos nas medidas durante as mudanças de correntes como quando a ação se expressam (13). Alguns amplificadores fornecem capacitância e "compensação" de resistência para a pipeta e vedação, o que limita a perda de sinal, mas estágios de alta resistência cabeça da maioria dos amplificadores impedem a compensação ideal. Em segundo lugar, uma situação artificial é imposta sobre a célula. Na tensão de clamp-mode, a área ao redor da membrana celular é mantido a um potencial fixo, nesses estudos, 0 mV. Isso não significa que não há corrente aplicada à membrana celular. O sinal medido em tensão de grampo é realmente a quantidade de corrente aplicada à membrana para manter o potencial fixo. Portanto, esta corrente aplicada pode alterar a atividade celular.
Gravações duplo no sistema de GnRH são particularmente difícil devido ao número limitado de neurônios GnRH e sua distribuição difusa. Para gravações de dupla para ser bem sucedido, o manipulador deve ser extremamente estável. Mesmo ligeiro movimento do eletrodo pode causar a pipeta para escorregar o neurônio e terminar a gravação. Além disso, o movimento da pipeta sobre a célula (por exemplo, re-posicionamento para compensar o movimento) pode alterar padrões de disparo. Alguns canais de íon, tais canais de cálcio tipo-N são mecanicamente sensíveis: trecho da membrana faz com que atividade repetitiva em configurações de gravação tanto de célula inteira e celular ligado (14). Finalmente, o sistema manipulador deve ser capaz de movimento extremamente fina e lisa. Como observado acima, com gravações dupla, um leva duas pipetas para a superfície dos dois neurônios previamente selecionados, ao mesmo tempo e as tentativas de fecharum neurônio. Se for bem sucedido, então as tentativas de fechar a segunda célula. Geralmente, não se pode esperar para selar e ter uma gravação de alta qualidade a cada tentativa. Isso, no entanto, cria um problema particular com gravações dual. Se um for bem sucedido com o primeiro neurônio, mas não com o segundo, é preciso mudar a pipeta e tentar uma célula diferente. Portanto, deve ser capaz de se mover tanto o objectivo de imersão do microscópio e da pipeta para o topo da perfusão bem (para alterar a pipeta) sem interromper o neurônio com sucesso selado.
Nosso desenvolvimento e uso da abordagem de células-anexado solta para gravações dupla é um grande avanço técnico em estudar neurônios GnRH. É provável produzir resultados úteis que irão ajudar a mover o campo para a frente, no contexto da questão crítica do que os mecanismos subjacentes à atividade coordenada que resulta em pulsátil secreção hormonal.
Acknowledgments
Sou grato a Ronald L. Calabrese, Jaeger Dieter (Emory University) e Yuhas Ward (Axon Instruments) para discussões técnicas úteis.
References
- Silverman, A. J. The gonadotropin releasing-hormone (GnRH) neuronal systems: immunocytochemistry. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Freeman, M. E. The neuroendocrine control of the ovarian cycle of the rat. In: The physiology of reproduction. The Physiology of Reproduction. Knobil, E., Neill, J. D. , Raven Press. New York. (1994).
- Belchetz, P. E., Plant, T. M., Nakai, Y., Keogh, E. J., Knobil, E. Hypophysial responses to continuous and intermittent delivery of hypopthalamic gonadotropin-releasing hormone. Science. 202, 631-633 (1978).
- Maurer, J. A., Wray, S. Luteinizing hormone-releasing hormone quantified in tissues and slice explant cultures of postnatal rat hypothalami. Endocrinology. 140, 791-799 (1999).
- Harris, G. C., Levine, J. E. Pubertal acceleration of pulsatile gonadotropin-releasing hormone release in male rats as revealed by microdialysis. Endocrinology. 14, 163-171 (2003).
- Sisk, C. L., Richardson, H. N., Chappell, P. E., Levine, J. E. In vivo gonadotropin-releasing hormone secretion in female rats during peripubertal development and on proestrus. Endocrinology. 142, 2929-2936 (2001).
- Cardenas, H., Ordog, T., O'Byrne, K. T., Knobil, E. Single unit components of the hypothalamic multiunit electrical activity associated with the central signal generator that directs the pulsatile secretion of gonadotropic hormones. Proc Natl Acad Sci U S A. 90, 9630-9634 (1993).
- Silverman, A. J., Wilson, R., Kesner, J. S., Knobil, E. Hypothalamic localization of multiunit electrical activity associated with pulsatile LH release in the rhesus monkey. Neuroendocrinology. 44, 168-171 (1986).
- Roberts, C. B., O'Boyle, M. P., Suter, K. J. Dendrites determine the contribution of after depolarization potentials (ADPs) to generation of repetitive action potentials in hypothalamic gonadotropin releasing-hormone (GnRH) neurons. J Comput Neurosci. 26, 39-53 (2009).
- Spergel, D. J., Kruth, U., Hanley, D. F., Sprengel, R., Seeburg, P. H. GABA- and glutamate-activated channels in green fluorescent protein-tagged gonadotropin-releasing hormone neurons in transgenic mice. J Neurosci. 19, 2037-2050 (1999).
- Nelson, J. F., Felicio, L. S., Randall, P. K., Sims, C., Finch, C. E. A longitudinal study of estrous cyclicity in aging C57BL/6J mice: I. Cycle frequency, length and vaginal cytology. Biol Reprod. 27, 327-339 (1982).
- Gee, D. M., Flurkey, K., Finch, C. E. Aging and the regulation of luteinizing hormone in C57BL/6J mice: impaired elevations after ovariectomy and spontaneous elevations at advanced ages. Biol Reprod. 28, 598-607 (1983).
- Anson, B. D., Roberts, W. M. Loose-patch voltage-clamp technique. Neuromethods, Patch-clamp analysis: Advanced techniques. Walz, W., Boulton, A. A., Baker, G. B. 35, Humana Press. Place Unknown. 265-286 (2002).
- Calabrese, B., Tabarean, T., Juranka, P., Morris, C. E. Mechanosensitivity of N-type calcium channel currents. Biophys J. 83, 2560-2574 (2002).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Rapid killing of single neurons by irradiation of intracellularly injected dye. Science. 206, 702-704 (1979).
- Higure, Y., Katayama, Y., Takeuchi, K., Ohtubo, Y., Yoshii, K. Lucifer Yellow slows voltage-gated Na+ current inactivation in a light-dependent manner in mice. J Physiol. 550, 159-167 (2003).

{kind=link}