1. Screening of Mutants with Altered Phototropic and Gravitropic Responses
- Prepare 1x Murashige and Skoog (MS) Medium, pH 5.7, with 0.8% Agar One Day Before the Experiment.
- Add 800 ml of double distilled water to a 2 L conical flask and stir with a magnetic bar.
- Add 4.4 g of MS salt to the conical flask.
- Add 0.5 g of 2-(N-Morpholino) ethane sulphonic acid (MES) and stir until all of the salts are completely dissolved.
- Adjust the pH of the medium to 5.7 with 1 M KOH.
- Transfer the medium to a mass cylinder and bring final volume to 1 L with ddH2O.
- Pour back the medium into a 2 L conical flask and add 8 g of plant agar.
- Wrap the mouth of the conical flask tightly with aluminum foil.
- Autoclave the MS medium and ddH2O for 15 min at 121 °C, 15 psi, and after autoclaving, keep the medium in a 60 °C incubator and the ddH2O at RT.
- After cooling to 60 °C, pour the medium into 125 x 125 x 20 mm3 square plates on a laminar flow bench (50 ml in each plate).
- Allow the agar to solidify at RT for about 1 hr by keeping the plates partially open. If the plates are not used immediately, seal them by using plastic wrap and store at 4 °C in a refrigerator.
- Surface-sterilize the seeds with 1.1% sodium hypochlorite and dot the seeds on 1x MS agar plates that were prepared in section 1.1.
Note: Here, Arabidopsis thaliana {Col-0 seeds, dsGSL8 (Col-0)}15 are used for tropic response, callose staining and HPTS loading. Arabidopsis thaliana {Col-0 and dsGSL8 (Col-0) EMS mutant} seeds (unpublished) are used for tropic response in Figure 2.- Autoclave 300 ml of ddH2O in a bottle.
- Add approximately one hundred seeds for each background, i.e., mutant/over-expression line and wild type (Col-0) seeds, to a 1.5 ml microcentrifuge tube.
- Take out 1x MS agar plates from a 4 °C refrigerator and dry them on a laminar flow bench in a partially open position.
- Surface-sterilize the seeds by adding 1 ml of a 0.2x bleach solution (1.1% sodium hypochlorite) and keep the microcentrifuge tubes on shaker at 250 rpm for 5 min.
- Let the seeds settle to the bottom by gravity and then remove the bleach solution by micropipette.
- Add 1 ml of fresh ddH2O (autoclaved) to the seeds and mix well by inverting the microcentrifuge tube several times. Let the seeds settle again and then remove the water.
- Repeat step 1.2.6 5-6 times.
- Add double the amount of water to the volume of the seeds that were left after the final washing.
- Dot the sterilized seeds on a semi-dried 1x MS agar plate using a 1 ml micropipette tip containing 200 µl of seeds (including water).
- Keep approximately a 0.1 cm distance between two seeds and a minimum of 1.8 cm between two seed rows. Dot the seeds in five horizontal rows per square plate (125 x 125 x 20 mm3). Allow the water to dry slightly before placing the lid onto the plate.
- Seal the plates with surgical tape, stack all of the plates vertically, and wrap with aluminum foil.
- Transfer the wrapped plates to a dark box (with no access to light). Then, keep the dark box in a 4 °C room/refrigerator for 3 days.
- Starting from this step, keep the direction of plates unchanged. After 3 days of cold treatment, keep the dark box in a plant culture room at 22 °C for the next 3 days. After 3 days, check the germination status of the seeds under green light in a dark room.
NOTE: Usually 3 day-old Col-0 etiolated seedlings can grow up to 1.6 cm in length.
- Analyze the altered phototropic and gravitropic response in mutants or over-expression lines of a specific gene. Select only similarly sized and vertically grown straight seedlings under green light in a dark room. Transfer selected seedlings to a fresh 1x MS agar plate using the tip of a sterilized toothpick.
- Gently select the seedlings from the lowest part of their hypocotyl without touching any other parts. Make sure that all of the seedlings have same cotyledon/hook orientation, and after transferring, that the orientation of seedling hooks remain the same. Use a minimum of 20 seedlings of each background in each row and a minimum of two plates for each time point for further analysis.
Note: Four different backgrounds can be compared in each plate (4 rows).
Optional: While using dexamethasone (dex)-inducible RNAi/over-expression lines, transfer the above seedlings to a 1x MS plate along with the +dex MS plate (plate containing 1x MS, 0.8% agar medium supplemented with 20 µM dex) for 3 hr and keep the plates vertically in dark box. - To check phototropic response, transfer the above plates vertically to a black box with only one side opened. Transfer the black box containing plates to a plant growth chamber with unilateral white light. To ensure unilateral light, place the edge of the plate toward the light source (not the front side). Refer to the setup in Figure 1.
- Similarly, to check the gravitropic response, keep the plates with transferred seedlings vertical, and cover with aluminum foil. Next, change the plate orientation vertically to 90° and keep it inside the black box covered from all sides. Refer to the setup in Figure 1.
- After the given time points, (usually 1.5 hr, 3 hr, 6 hr, and 12 hr) scan the plates with a scanner, and save the pictures in JPEG format.
- Measure the bending angle of seedlings using ImageJ software (http://imagej.nih.gov/ij/index.html) (Figure 2).
- Start the ImageJ program by double clicking (Figure 2A).
Note: The major tools that were used in the angle measurement are shown in Figure 2B. - Click the "file" option followed by sub-option "open" in the ImageJ toolbar, and open pictures saved in JPEG file format to measure the tropic bending angle (Figure 2C).
- Click the magnifying tool to zoom in or out on the picture by clicking the left or right mouse button, respectively as shown in Figure 2B.
- Click the scrolling tool to drag and position the picture (Figure 2D).Next, click the angle tool to measure the angle of bending (Figure 2E).
- Make a double left click on the original hypocotyl growing direction point 'a', drag the mouse to the bending initiation point 'b' (Figure 2F), and click left button to draw the first line (Figure 2G).
- Drag the mouse from point b to the bending end point 'c', and click the left button to draw the second line (Figure 2H) and form a fixed
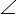 A. Note: The true bending angle is B, which is equal to 180°-A. (Figure 2F).
A. Note: The true bending angle is B, which is equal to 180°-A. (Figure 2F). - Measure and record A by pressing 'M' on the keyboard. The result file will open separately as shown in Figure 2I. Measure and record all of the seedlings one by one. For example, first measure the bending angle for all of the hypocotyls from Col-0 and then for mutant lines.
- Start the ImageJ program by double clicking (Figure 2A).
- Transfer the dataset from ImageJ to spreadsheet file to make the average of bending angles of each background.
- For quantitative analysis, make a bar graph diagram with statistical test results, including standard error bars and probability values (See Figure 2).
- Calculate the true bending B, and design a formula using spreadsheet, i.e., B = 180°-A. Note: A is the value that was derived from ImageJ analysis.
- Measure the average bending angle B for each background using spreadsheet tools (Figure 2J).
- Analyze the standard deviation between replicates for each measurement using the spreadsheet "formulas" tool (Figure 2J).
- Test the significance of the results by applying a t-test to the average bending angle and standard deviation values calculated as mentioned above.
- Draw a bar diagram using the above values to graphically represent the quantitative difference in the bending angle between two different backgrounds using the spreadsheet "insert" tool (Figure 2K).
Note: Mutant/over-expression lines of strong candidate genes would show clear differences with respect to wild type, such as no bending, fast bending or slow bending.
- Calculate the true bending
- Gently select the seedlings from the lowest part of their hypocotyl without touching any other parts. Make sure that all of the seedlings have same cotyledon/hook orientation, and after transferring, that the orientation of seedling hooks remain the same. Use a minimum of 20 seedlings of each background in each row and a minimum of two plates for each time point for further analysis.
2. Screen Plant Lines with Defective Tropic Responses Due to Changes in PD SEL with an Altered PD Callose Level
- Perform a Hypocotyl Loading Assay by 8-hydroxypyrene-1,3,6-trisulfonic acid (HPTS) Dye Loading on the Top of Seedlings with HPTS Agarose Blocks and Compare the Movement of Dye Between Mutant/Over-expression Lines and Col-0.
- Prepare HPTS agarose blocks for the hypocotyl loading assay.
- Take 10 ml of ddH2O in a 50 ml conical flask and add 0.1 g of agarose to prepare a 1% agarose gel. Boil the agarose in a microwave oven for 1-2 min with intermittent shaking, and allow it to cool for 5 min.
- Add 50 mg of HPTS to 10 ml of 1% agarose solution, and mix well until the solution turns green.
- Pour the above mixture to a 10 mm Petri dish, and allow it to solidify for 15 min.
- Cut the solidified HPTS agarose with a sharp dissection blade to make small agarose blocks (0.5 x 2 cm2).
- Prepare plant samples for the HPTS dye-loading assay.
- Set a platform for the HPTS dye loading assay; place a microscope cover slide (24 x 50 mm2) on a new MS medium plate as shown in Figure 3A.
- Excise 3 day-old etiolated seedlings from the base of the hook using a sharp surgical scissor.
- Immediately transfer the above seedlings (a minimum of 5 seedlings from each background) to the surface of a microscopic cover glass in such a way that only 0.5 cm of hypocotyl from the excision position should remain on cover slide, and the rest of the hypocotyl should touch the medium as shown in Figure 3B.
- Place the HPTS agarose block on top of the hypocotyl (with an excised hook) for 5 min in such a way that the excised area touches the HPTS agarose block.
- After 5 min of loading, remove the agarose block from the hypocotyl surface, transfer the hypocotyls to the 10 mm Petri dish containing ddH2O and wash hypocotyls in ddH2O for 15 min with continuous shaking.
- Prepare Samples for Confocal Microscopy.
- Select a minimum of 5 seedlings from each background for observation under the confocal laser scanning microscope.
- In the laser channel, set the wavelength to 488 nm for specific excitation and 500 to 550 nm for band-pass emission.
- Transfer seedlings in 2 sets (one Col-0 + one mutant) to a new microscopic slide after washing with ddH2O. Add 100-150 µl of double-distilled water to the slide, and cover with a cover slide. Shift the slide to the stage of the confocal microscope.
- Focus the hypocotyl region (with a 20X or 40X objective lens) using bright-field (white light) illumination. Then, scan the slides for fluorescence.
- Take images for a minimum of 10 sets, and compare the two backgrounds by checking the extent of dye movement from the point of loading.
Note: Mutant/over-expression lines with altered symplasmic movement will show different patterns of dye loading compared to wild type Col-0.
- Prepare HPTS agarose blocks for the hypocotyl loading assay.
- Analyze the Callose Level in Mutant/Over-expression Lines Showing Altered Phototropic and Gravitropic Response and Showing Distinct HPTS Dye Movement Compared to Col-0.
- Prepare aniline blue callose staining dye.
- Make 1 M glycine, pH 9.5 and 0.1% (W/V) aniline blue in autoclaved ddH2O.
- Make the staining buffer by mixing 0.1% aniline blue and 1 M glycine in 2:3 volume ratios. For example, for 50 ml of staining buffer, take 20 ml of 0.1% aniline blue and 30 ml of 1 M glycine. Note: Make the staining buffer a minimum of 48 hr prior to use and store it in the dark.
- Stain bended or non-bended seedlings with callose-specific aniline blue dye.
- To inhibit the de novo callose synthesis during the staining process, add the final volume of 1.5 mM 2-deoxy-D-glucose (DDG) to 50 ml of staining buffer.
- Take 3-day-old etiolated seedlings for callose staining (with or without tropic response).
- Cut the seedlings from their hook region using thin surgical scissors.
- Transfer the seedlings in a small Petri dish containing 2 ml of staining buffer using the tip of a toothpick.
- Incubate the seedlings with staining buffer in the dark for 2 hr with continuous shaking at 30 rpm.
- After staining, wash the seedlings with ddH2O for 2 min to remove excess staining buffer.
- Select a minimum of 5 seedlings for each plant background for observation under the confocal laser scanning microscope.
- Prepare samples for confocal microscopy:
- Transfer the seedlings in set (one Col-0 + one mutant) to clean the microscope slides after washing with ddH2O.
- Add 100-150 µl of ddH2O and cover with a cover slide. Shift the slide to the stage of the confocal microscope.
- In a laser channel, set the wavelength to 405 nm for specific excitation and 500 to 550 nm emission for the imaging of aniline blue fluorescence.
- Bring the hypocotyl region into focus, first with 10X and later with 20X or 40X objective lenses using bright-field (white light) illumination.
- Then, switch on filtered light, scan the slides for fluorescence, and save the images.
- Measure the callose signal intensity with ImageJ software (http://imagej.nih.gov/ij/index.html) (See Figure 4).
- Change the format of confocal microscopy pictures from Olympus Image Binary format (*.oib) to JPEG file format. Note: Only JPEG format is supported by ImageJ software.
- Start the ImageJ program by double clicking (Figure 4A). Note: Arrows indicate the rectangle tool to be used in the callose intensity measurement (Figure 4B).
- Click the "file" option followed by the sub-option "open" in the ImageJ toolbar, and open microscopy pictures that were saved in JPEG file format to measure the callose intensity (Figure 4C).
- Click the rectangular icon on the ImageJ toolbar (Figure 4D).
- To analyze the signal intensity, drag the cursor over the picture and select the area to be examined (Figure 4E)
- Measure and record the numerical values for signal intensity by pressing 'M' on the keyboard (Figure 4E). Note: The result file will open separately (Figure 4F).
- Measure and record the intensity values one by one for all of the seedlings from each plant background.
Note: In ImageJ, the result file "Mean" indicates the signal intensity of the selected "Area".
- Copy the mean values from the ImageJ result file, and paste in spreadsheet file for quantitative analysis.
- For quantitative analysis, make a bar graph diagram with statistical tests, including standard error bars and probability values (Figure 4).
- Measure the average signal intensity (mean from ImageJ) for each background using the spreadsheet "formulas" tool (Figure 4G).
- Analyze the standard deviation between replicates for each measurement using the spreadsheet "formulas" tool (Figure 4G).
- Calculate the standard error (SE) using thespreadsheet "formulas" tool.
Note: SE = s/√n, where (s) is the standard deviation of the population, and (n) is the size (number of observations) of the sample. - Test the significance of the results by applying a t-test to the average signal intensity and standard error values calculated as mentioned above.
- Draw a bar graph diagram using the above values to graphically represent the quantitative difference in the callose level between two different backgrounds using the spreadsheet "insert" tool (Figure 4H).
Note: Mutant/over-expression lines of strong candidates will show different levels of callose compared to wild type, such as no callose, little callose or hyper PD callose accumulation.
- Prepare aniline blue callose staining dye.
In the current setup, dexamethasone (dex)-inducible RNAi lines of AtGSL8 [hereafter dsGSL8 (+dex/-dex)] were used, as homozygous gsl8 T-DNA insertion mutants are seedling lethal18. Three-day-old etiolated seedlings of dsGSL8 and wild type seedlings with ±dex were exposed to phototropic and gravitropic stimuli. We found that dsGSL8 (+dex) seedlings were defective in phototropism and gravitropism15. Figure 5 clearly shows that dsGSL8 (+dex) exhibits no bending under the influence of both phototropism and gravitropism. Furthermore, alteration in symplasmic movement in dsGSL8 (+dex) and dsGSL8 (-dex) or Col-0 was analyzed by a HPTS loading assay. Consistent with our previous finding, the HPTS dye movement in dsGSL8 (+dex) was substantially more extensive than in dsGSL8 (-dex) or Col-0, as shown in Figure 6. Callose is one of the key regulators of PD SEL, and AtGSL8 is known as a PD callose synthase16. Furthermore, callose aniline blue staining was carried out, and dsGSL8 (+dex) have persistently shown a low PD callose level before and after tropic responses, as shown in Figure 7.

Figure 1. Flow chart of the steps that are involved in the stimulation of phototropic and gravitropic responses. (A) Transfer vertically grown seedlings from different plant backgrounds to one plate but in separate lines. (B) For phototropism, keep the plates facing towards unilateral white light in a dark box that is opened on one side. For gravitropism, first wrap the plates with aluminum foil, rotate 90°, and transfer to a dark box (covered from all sides). (C) Using a scanner, scan the plates after different intervals of time, such as 1.5 hr, 3 hr, 6 hr and 12 hr. (D and E) These panels show data generation from ImageJ and data analysis using spreadsheet files, respectively. Please click here to view a larger version of this figure.

Figure 2. Measurement of the bending angle by ImageJ. (A) View the Image J program by double clicking. (B) The major tools that were used in the angle measurement. (C) Open the "File" option followed by the sub-option "open" in the Image J toolbar to open microscopy pictures that were saved in JPEG file format. (D) Use the magnifying tool to zoom in or out on the picture by clicking the left or right mouse button, respectively. (E) Use the angle tool to measure the angle of bending. (F) Outline of the angle measurement: abc defines a fixed A, and the true bending angle is calculated as B, which is equal to 180°-A. (G) Line representing the distance from point a to point b. (H) Continuity of line a-b to point c making A. (I) Result file of Image J showing A value. (J) Representative spreadsheet file displaying the average bending angle measurements, standard deviations, standard errors and t-test calculations. (K) Bar graph diagram displaying the bending angle measurements between two different plant backgrounds Col-0 and dsGSL8 EMS mutant. Error bars are SEM (standard error of the mean +) and *** represents the significance by T-test (p value >0.001). Please click here to view a larger version of this figure.

Figure 3. Outline of the steps that are involved in the HPTS loading assay. (A) Panel showing the preparation of HPTS agarose blocks. (B) Panel representing the excision and transfer of seedlings and the loading of HPTS agarose blocks. (C) Panel displaying the steps that are involved in sample preparation for confocal imaging. Please click here to view a larger version of this figure.

Figure 4. Measurement of the callose intensity by ImageJ. (A) View the ImageJ program by double clicking. (B) The major tools that were used in the callose intensity measurement. (C) Open the "File" option followed by the sub-option "open" in the ImageJ toolbar to open microscopy pictures that were saved in JPEG file format. (D) The rectangular icon on ImageJ toolbar. (E) The selected area of the image for analyzing the callose intensity. (F) ImageJ result file displaying the callose intensity value as "Mean". (G) Representative spreadsheet file displaying the average callose intensity measurements, standard deviations, standard errors and t-test calculations. (H) Bar graph diagram representing the callose intensity difference between the two plant backgrounds. Error bars are SEM (standard error of the mean +) and *** represents the significance by T-test (p value >0.001). Please click here to view a larger version of this figure.

Figure 5. Reduction of AtGSL8 expression leads to defective tropic responses. (A) Phototropic response showed by dsGSL8 (±dex) along with Col-0. (B) Gravitropic response shown by dsGSL8 (±dex) along with Col-0. dsGSL8 (+dex) shows no phototropic or gravitropic responses. dsGSL8 (+dex) and dsGSL8 (-dex) symbolize dexamethasone-treated and untreated dsGSL8-inducible RNAi lines, respectively. Scale bar represents 0.2 cm. Please click here to view a larger version of this figure.

Figure 6. The symplasmic movement increased after the suppression of AtGSL8. Fluorescence images were taken after HPTS loading to dsGSL8±dex hypocotyl cut surfaces along with wild type Col-0. dsGSL8 (+dex) showed more extensive movement of HPTS dye, demonstrating increased symplasmic movement. Scale bar represents 0.2 mm. Please click here to view a larger version of this figure.

Figure 7. AtGSL8 suppressed lines showed symmetric PD callose distribution between illuminated and shaded sides of the hypocotyl. (A) Fluorescence image showing aniline blue callose staining for dsGSL8 (-dex) at 0 hr. (B) Fluorescence image showing aniline blue callose staining for dsGSL8 (-dex) after 6 hr of phototropism. (C) Fluorescence image showing aniline blue callose staining for dsGSL8 (+dex) after 6 hr of phototropism. Yellow arrows indicate the callose accumulation on shaded region of hypocotyl. Scale bar represents 50 µm. (D) Bar graph diagram representing the amount of PD callose that accumulated in dsGSL8 (±dex) hypocotyls before and after 3 hr and 6 hr of phototropism. Fluorescence foci intensities were measured from ten independent hypocotyls (data are mean ± SD; Student's t-test, *p <0.01). This figure has been modified from (Han et al., 2014) 15. Please click here to view a larger version of this figure.
