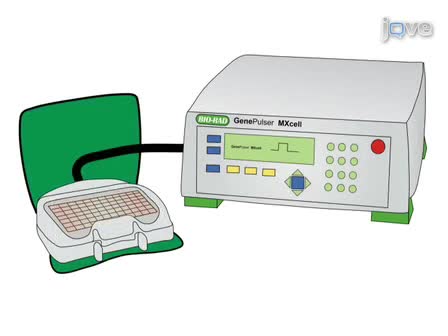
Osteoclasts are the principal bone-resorbing cell in the body. An ability to isolate osteoclasts in large numbers has resulted in significant advances in the understanding of osteoclast biology. In this protocol, we describe a method for isolation, cultivating and quantifying osteoclast activity in vitro.