

Summary
Een krachtige manier om de neuronale lawines, dat wil zeggen schaal-invariant spatio-temporele activiteit uitbarstingen, indicatief van kritieke toestand dynamiek in cortex te bestuderen. Lawines ontstaan spontaan in de ontwikkeling van oppervlakkige lagen van gekweekte hersenschors, die voor de lange termijn metingen van de activiteit mogelijk maakt met vlakke geïntegreerde multi-elektrode arrays (MEA) onder nauwkeurig gecontroleerde omstandigheden.
Abstract
De cortex is spontaan actief is, zelfs bij het ontbreken van een bepaalde input of motor output. Tijdens de ontwikkeling, deze activiteit is belangrijk voor de migratie en differentiatie van cortex celtypen en de vorming van neuronale verbindingen 1. In het volwassen dier, lopende activiteiten weerspiegelt het verleden en de huidige stand van een dier, waarin zintuiglijke prikkels naadloos zijn geïntegreerd in toekomstige maatregelen te berekenen. Dus, een duidelijk inzicht in de organisatie van de lopende ie spontane activiteit is een voorwaarde om cortex functie te begrijpen.
Tal van opnametechnieken blijkt dat de lopende activiteit in cortex bestaat uit vele neuronen waarvan de afzonderlijke activiteiten tijdelijk bedrag grotere evenementen die in de lokale veld potentieel (LFP) worden gedetecteerd met extracellulaire micro-elektroden, of in het elektro-encefalogram (EEG), de magnetoencephalogram (MEG ), en het BOLD signaal van functionele magnetische resonantie imaging (fMRI). De LFP is op dit moment de methode van keuze bij het bestuderen van neuronale activiteit de bevolking met een hoge temporele en ruimtelijke resolutie bij de mesoscopische schaal (enkele duizenden neuronen). Op de extracellulaire micro-elektrode, lokaal gesynchroniseerd activiteiten van ruimtelijk grensde neuronen leiden tot snelle ombuigingen in de LFP tot enkele honderden microvolt. Bij gebruik van een scala aan micro-elektroden, kunnen de organisaties van dergelijke verlegging gemakkelijk worden bewaakt in ruimte en tijd.
Neuronale lawines beschrijven de schaal-invariant spatiotemporele organisatie van de lopende neuronale activiteit in de hersenen 2,3. Ze zijn specifiek voor de oppervlakkige lagen van de cortex, zoals vastgesteld in vitro 4,5, in vivo in de verdoofde rat 6, en in de wakkere aap 7. Belangrijk is dat zowel de theoretische en empirische studies 2,8-10 suggereren dat neuronale lawines een prachtig gebalanceerde kritieke toestand dynamiek van de cortex dat de informatie-overdracht en verwerking van gegevens optimaliseert aan te geven.
Om de mechanismen van de neuronale lawine ontwikkeling, onderhoud, studie-en regelgeving, in vitro voorbereidingen zijn zeer nuttig, omdat ze zorgen voor een stabiele opnames van de lawine activiteit onder nauwkeurig gecontroleerde omstandigheden. Het huidige protocol wordt beschreven hoe u neuronale lawines in vitro onderzoek door gebruik te maken van de oppervlakkige laag ontwikkeling in organotypische cortex culturen, dat wil zeggen slice culturen, geteeld op vlakke, geïntegreerde micro-elektrode arrays (MEA, zie ook 11-14).
Protocol
1. Steriel, afsluitbare glazen kamer met MEA voor de lange termijn Recordings
- Schroefdraad glazen cilinders met Teflon-plastic kap (Ace Glass), die nodig zijn voor veilige en strakke cultuur kamer sluiting, worden gesneden (Aceglass) ongeveer 2 mm van de onderkant van de draad (Fig. 1A, B). Schoon glas ringen door te spoelen met water (3x) en koken gedurende 5 minuten in 200 proof ethyl-alcohol, laten drogen.
- Aliquot silicium-oplossing nodig is om glazen ringen hechten aan MEA oppervlak. Meng 15 ml van de delen A & B van Sylgard 184 Silicone elastomeer Kit grondig, laat zitten voor 15 min om luchtbellen te verwijderen, op te slaan in 1 ml porties bij -20 ° C.
- Lijm glas ring om de MEA (8x8 raster w / interne grond-elektrode, 30 urn elektrode diameter van 200/100 um inter-elektrode afstand voor rat / muis) (fig. 1A, B). Neem 1 ml van silicium (23 ° C) in spuit met een kleine naald. Van toepassing silicone om ongepolijste snijvlak van glas ring, in het midden glazen ring op MEA van toepassing een extra laag van silicium rond de buitenkant van de ring voor een sterkere seal, laat remedie voor 1 - 2 uur bij ~ 60 ° C op hete plaat.
- Steriliseren MEA kamer en kamer caps in een laminaire stroming kap door 3x spoelen in gedemineraliseerd water, gevolgd door 70% alcohol (3 x, voor de laatste spoeling te laten zitten voor 10 min in alcohol), gevolgd door 10 minuten blootstelling van kamer en kap het interieur tegen UV-licht . Autoclaaf MEA kamer (120 ° C; nat; 45 min) en laat drogen.
- Vacht MEA oppervlak in cultuur kamer met poly-D-lysine. Voor nieuwe MMO's, die eerder lipofiel, jas door herhaalde druppel aspiratie van de oplossing van de elektrode net. Voor gebruikt MMO's, kamer bodem met oplossing te dekken, aspiratie overtollige vloeistof, laten verdampen onder steriele omstandigheden binnen laminaire capuchon. Bevestig de dop om MEA kamer afdichting voor de opslag en het toekomstige gebruik.
2. Ingrediënten die nodig zijn voor de voorbereiding en de groei van organotypische Culturen
- Los steriele agar in 0,9% NaCl, giet het in steriele petrischaal (Falcon, 100 x15; ~ 5 mm-niveau), laat afkoelen, en steriele wrap met Parafilm voor opslag. Snijd 20 x 10 x 5 mm blokken van massief agar voor gebruik.
- Store superlijm, bijvoorbeeld Devcon Super Glue II, met de verpakking afgeveegd neer met 70% EtOH voor het openen, in de laminaire flow kap de steriliteit te behouden.
- Bereid 50% D-Glucose (SIGMA ultra, G7528), door het toevoegen van 40 g glucose tot 40 ml gedeïoniseerd water cultuur (Sigma). Bewaren in 2 ml bij -20 ° C.
- Voeg 4 ml van 50% D-glucose tot 500 ml van de Balanced Salt oplossing Gey, en chill om slush (mengsel van vloeibare / ijskristallen) in de vriezer voor gebruik.
- Reconstrueren kip plasma in 5 ml gedeïoniseerd water cultuur (schud zacht, te voorkomen de vorming van luchtbellen), laat-oplossing rusten gedurende 5 - 10 min, zachtjes swirl, en decantate de heldere inhoud in een steriele petrischaal. Steriel-filter (0,22 um porie-filter; eiwit) plasma-oplossing, hoeveelheid 350 pl in cryotubes (NuncTM), op te slaan bij -20 ° C.
- Reconstitueer trombine van runderen plasma dienovereenkomstig, steriel-filter (0,22 um porie filter), 40 pi aliquot in cryotubes (NuncTM), op te slaan bij -20 ° C. Voor de werkende oplossing, verdund 40 ul van de trombine-oplossing in 375 ul van gebalanceerde zoutoplossing Gey's W / D-glucose.
- Bereid 400 ml kweekmedium door het mengen van 100 ml van het paard serum, 200 ml basismedium Eagle, 100 ml gebufferde fysiologische zoutoplossing Hank's oplossing voor die 4 ml 50% glucose en 2 ml van 200 mM L-glutamine toegevoegd. Kunnen worden opgeslagen 4 - 8 weken in 100 ml PYREX flessen bij 4 ° C.
- Bereid mitose remmer door het mengen van 0,3 mM uridine, 0,3 mM ARA-C cytosine-β-D-arabinofuranoside, en 0,3 mM 5-fluoro-2'-deoxyuridine, steriel filter, aliquot 200 ul en bewaar bij -20 ° C voor 6 - 12 maanden.
3. Cortex en ventrale tegmentale gebied (VTA) Tissue Dissection (tijd: <1 uur)
- Procedure opbrengst cortex en de VTA weefsel voor ~ 12 co-culturen van ratten of muizen, en is bereid in een laminaire stroming kap onder steriele omstandigheden. Totale tijd voor het verzamelen van weefsel moet worden dan een uur.
- Neem gezond, goed gevoed pups (worpgrootte ~ 10, de aanwezigheid van een abdominale 'melk spot') op 1-2 postnatale dag (PND). Voorzichtig Houd een pup van de snuit, laat het dan vrij te hangen, en snel onthoofden aan de basis van de nek met een scherpe schaar.
- Voor de hersenen verwijderen, verwijder de huid (twee zijdelingse schaar sneden), knippen schedel open met veel oog schaar (een sagittale middellijn gesneden; een coronale afgesneden cortex / cerebellum kruising). Flip terug alle 4 schedel flappen. Met een scherpe spatel, frontaal dwars door de bulbus olfactorius, vooraf spatel caudaal onder de hersenen. Til de hersenen uit de schedel en laat hem glijden in steriele, gekoeld, oplossing Gey's voor snel koelen en tijdelijke opslag. Herhaal de stappen 3,2 tot 3,3 voor 2 meer hersenen (totale tijd: <20 min).
- Voor het verkrijgen van VTAweefsel, overdracht van de hersenen op een steriel, droge petrischaal met een kleine spatel. Verder te verwijderen overtollige vloeistof door zachtjes te schuiven elke hersenen ongeveer 1 cm opzij. Verwijder de hersenstam door een coronale, verticale snede op het niveau van het cerebellum met behulp van een scheermesje.
- Lijm agar blok op schijf voor montage mechanische stabilisatie van de hersenen tijdens het snijden procedure. Leg een dunne lijn van de secondenlijm een paar millimeter in de voorkant van de agar blok op de schijf (vermijd lijm aanraken van de agar).
- Met behulp van een kleine spatel, de overdracht en de bevestiging per hersenen, frontale paal naar beneden. Zorg ervoor dat de frontale polen zijn gelijmd montage schijf en dat ventrale zijden van de agar aanraken zonder lijmresten om een goede mechanische stabilisatie bereiken tijdens het snijden en gemakkelijk lift-off van gesneden plakjes.
- Voorzichtig onderdompelen en veilig montageschijf met hersenen montage in een vibratome tray (bijv. Leica VT1000) gevuld met een steriele, gekoeld oplossing Gey's. Met een zorgvuldig gereinigd scheermesje (90% EtOH), snijd coronale plakjes van de middenhersenen in de hoogste trillingsfrequentie en de relatief lage voorwaartse snelheid bij een dikte van 400 500 pm. Met behulp van een omgekeerde Pasteur pipet met afzuiging lamp, de overdracht en het verzamelen van plakjes met de VTA in 35 x 10 mm petrischalen gevuld met een steriele, gekoeld oplossing Gey's (Fig. 1C, zie ook coronale plaat 18 tot 20 op E22 in 15).
- Voor de cortex secties, herhaalt u de stappen 3,2 tot 3,6, maar verticale snede tussen de cortex en het cerebellum, toepassen en monteren forebrains met frontaal paal boven. Ongeveer 3 coronale slices (350 micrometer dikte) te beginnen op het niveau van de striatum worden verzameld voor de toekomst cortex dissectie.
- Met behulp van een micro-mes van gebroken scheermesjes, ontleden ~ 2 mm breed coronale deel van frontale cortex en de middenhersenen gebieden met de VTA (Fig. 1C) onder een stereomicroscoop. Gescheiden in te zamelen weefselcoupes in kleine gerechten (bv. kamer dia's) gevuld met de oplossing gekoeld Gey's.
4. Montage Cortex en VTA Tissue Slices op de MEA (tijd: <1 uur)
- Positie MEA bij kamertemperatuur onder een stereo microscoop met de elektrode array in focus. Center een 25 ul druppel van plasma op de schone, stof-vrij, en steriele elektrode-array matrix. Met behulp van kleine spatulae, schuif een cortex en VTA sectie in het plasma druppel.
- Plaats MEA op koelplaat, heroriëntatie bekijken, laat chill voor ~ 15 s, voeg dan 25 ul van trombine in het plasma druppel. Met behulp van de trombine pipetpunt, zorgvuldig verspreid de plasma / trombine mengsel met kleine cirkelvormige bewegingen over de MEA. Raak de broze elektrode array. Voorzichtig de positie van de cortex op de array met dorsale rand langs de tweede elektrode rij van de array. Op deze manier zal de ontwikkeling van oppervlakkige lagen uiteindelijk betrekking hebben op de herinnering van de array. De VTA is naast geplaatst om de ventrale rand van de cortex sectie (fig. 1D).
- Cap en losjes sluit de MEA kamer aan hoge vochtigheid te behouden, terwijl de MEA / cultuur montage zit voor ~ 5 min in de kap op kamertemperatuur te laten voor plasma / trombine coagulatie. Ondertussen, herhaal dan stap 4,1 tot 4,3 voor 3 meer culturen.
- Voeg voorzichtig 600 ul kweekmedium in kleine druppeltjes aan de cultuur kamer met een 1 cc spuit met 25 x 5 / 8 naald.
- Goed afsluiten van de MEA kamer en plaats MEA / cultuur montage op de schommelende opbergvak in de couveuse (Fig. 1B). Het versnellen van de procedure, 3 - 4 MMO's kunnen worden gemonteerd in overlappende sequenties. Montage tijd voor 12 MMO moet worden <1 uur.
- Na 2 dagen in vitro (DIV), voeg 10 ul van mitose-remmer. Refresh cultuur media met 60% op 4 DIV en om de 4 dagen daarna.
5. Elektrofysiologische opname en Stimulus Generation
- Om de relatie tussen belangrijke ombuigingen in de lokale veld potentieel (LFP), en de neiging van neuronen tot actie potentialen brand vast te stellen, na ~ 1 week 5,6 opnemen spontane activiteit bij 24 kHz voor ~ 10 min van elke elektrode van de MEA (Hardware : MEA1060 w / blanking circuit, x 1200 te krijgen, 12 bit A / D, het bereik 0-4096 mV, MultiChannel Systems, Software: MC_Rack). De grond wordt verleend via de interne massa-elektrode, of extern door het toevoegen van een Ag / AgCl-half cel.
- Scheid de LFP met een band pass filter van 1 - 200 Hz van extracellulaire spike activiteit (band pass 300 - 3000 Hz). Spike activiteit kan verder worden onderverdeeld in single-en multi-eenheid activiteit met behulp van off-line spike sorteerders (bijv. Plexon Inc.) Bereken spike geactiveerd gemiddelden voor elke elektrode. Voor de cortex culturen, zullen de meeste gemiddelden te identificeren negatieve LFP doorbuigingen (nLFP) als de gewenste tijd van neuronale pieken in de cultuur.
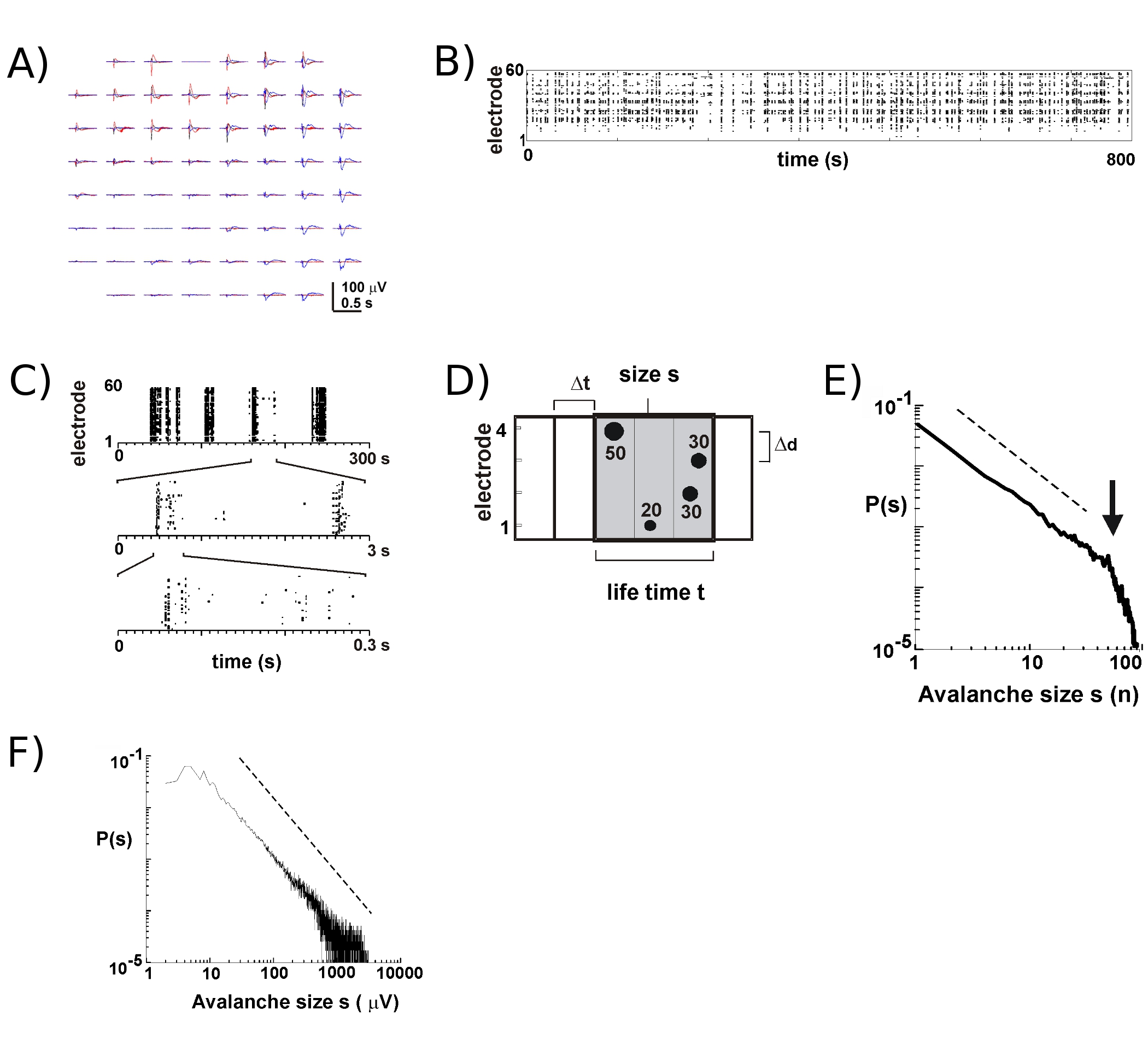
- Bereken voor elke elektrode een drempel van -3 standaarddeviaties van het geluid (SD) van de LFP sporen (fig. 2), Bepalen de piekuren en de amplitudes van nLFPs dat kruis drempel (Fig. 2B, C). Kies een tijd bin At (bijv. tussen 2 - 8 ms) en identificeer spatiotemporele nLFP clusters op de array door de aaneenschakeling van nLFPs van alle elektrode die in dezelfde tijd van opeenvolgende combineert met een lengte At (fig. 2D, voor details zie 2,4 , 5).
- Met het oog op neuronale lawines te identificeren, het berekenen van de grootte van elk nLFP cluster, bijv. het aantal actieve elektroden of de som van nLFP amplitudes, de bouw van een groot histogram, en plot in dubbel-logaritmische coördinaten. Voor de neuronale lawines, de grootteverdeling volgt een machtswet benaderd door een rechte lijn in dubbel-logaritmische coördinaten 2 (fig. 2E, F). Zie 16 voor statistische tests op de macht wetten.
- Uitlokken opgeroepen reacties in het weefsel door het selecteren van een elektrode, waardoor de huidige gecontroleerde stimuli met een amplitude S zijn toegepast (Stimulus generator STG 1008, Multichannel Systems). Te verminderen elektrode schade, gebruik maken van een assortiment beperkt, laad-neutraal stimulatie van enkele schokken met een bipolaire blokgolf: 50 microseconden met amplitude-S, gevolgd door 100 microseconden met amplitude + S / 2 en S tussen de 10 tot 200 uA. Zie de handleiding voor meer informatie.
- Voor het opnemen van het dynamisch bereik 9, record stimulus opgenomen LFP reacties op 4 kHz sampling rate op alle elektroden na 500 ms na stimulatie. Gebruik blanking circuit (Multi-channel systemen) waarop het hoofd podium versterkers verbreekt tijdens de stimulatie om stimulus artefacten te verminderen en de voorversterker verzadiging te voorkomen.
6. Representatieve resultaten:
Met de nieuwe MMO's ongeveer 8 - 9 van 10 culturen zullen overleven voor vele weken. De meeste van onze lange-termijn-opnamen vinden plaats in de couveuse in de cultuur medium, die ons in staat om de ontwikkeling van individuele culturen te volgen in de loop van vele weken 5. Op basis van onze experimenten, kan LFP opnames betrouwbaar worden verkregen met MEA gebruikt voor meer dan 100 dagen cultuur. In tegenstelling, extracellulaire spike activiteit is meer betrouwbaar gemeten worden met relatief nieuwe MEA (<40 dagen cultuur). In een typisch experiment, we dragen een MEA van de opslag lade (Fig. 1B, rechts) om de lade met het hoofd podium geplaatst (Fig. 1B, links) het bijhouden van de cultuur kamer verzegeld. Voor de cortex vijf, cortex-VTA co-culturen 6, alsmede in de verdoofde rat 6 en de wakkere aap in vivo 7, neuronale tijdens neuronale lawines in oppervlakkige lagen komt voornamelijk dicht bij de top negatieve uitslag van de LFP (nLFP ). Zo kan de spatiotemporele organisatie van lokaal gesynchroniseerde neuronale groepen worden geschat door het meten van het optreden van nLFPs in ruimte en tijd op de array 17.
Activiteit op de MEA neigt te ontstaan in tijdelijke clusters, is zodanig dat de activiteit op een elektrode gepaard gaan met activiteiten op andere sites. Typische golfvormen van de LFP tijdens dergelijke activiteit periodes zijn weergegeven in figuur 2A, door meer dan plotten drie clusters die zich enkele seconden uit elkaar. Voor elke cluster, kunnen negatieve veld doorbuigingen te zien op verschillende elektroden binnen een venster van een s. Bij het uitpakken nLFP pieken die een drempel van meerdere negatieve SD kruis, de activiteit in de vorm van nLFP piektijden is gunstig gevisualiseerd in een raster waarin de 'kolommen' van de stippen in de buurt van samenvallende nLFPs op verschillende elektroden (Fig. 2B). De spatiotemporele organisatie van deze activiteit is tamelijk complex; 'zuilen', die lijken min of meer homogene lage temporele resolutie, zijn samengesteld uit verschillende clusters bij hogere temporele resolutie en ga zo maar door (fig. 2C). In feite is de opkomst van spatiotemporele nLFP clusters zeer georganiseerd in corticale netwerken. Meer specifiek, de organisatie is schaal-invariant voor neuronale lawines. Dit wordt aangetoond door het berekenen van de kans op clustergrootten bij een bepaalde temporele resolutie At. Hier worden clusters bestaan uit nLFPs die zich voordoen in dezelfde of opeenvolgende keer bakken (fig. 2D). Wanneer de omvang van een dergelijke cluster wordt uitgedrukt in het totale aantal nLFPs per cluster, of geïntegreerd nLFP amplitudes per cluster, de clustergrootte distributies onthult een power wet, waarvan de helling heeft aangetoond dat -1,5 2,4,5,7 ( Fig. 2E, F). Merk op dat deze verdeling een schaal-invariant bestellen van clustergrootten dat is de verhouding van de maten S tot kxs, waarbij k een constante factor identificeert, is k -1,5, die onafhankelijk is van s. Deze bevoegdheid wet organisatie is onafhankelijk van de grootte van de array 2, van de temporele resolutie At 2, en de drempel wordt gebruikt om belangrijke nLFP identificeren doorbuiging 7. Omdat nLFP amplitude schalen met neuronale grootte van de groep 7, de schaal-invariante organisatie van nLFPs weerspiegelt een schaal-invariant, dwz fractal, bestellen of lokaal gesynchroniseerd neuronale groepen die alle maten op te nemen.

Figuur 1. (A) Side en top uitzicht op de MEA met schroefdraad glas ring gemonteerd, en de bijbehorende cap. (B) Binnenkant van de incubator. Links: headstage mount waardoor het opnemen van een enkele cultuur onder incubator voorwaarde. Rechts: Lade holding tal van MMO's voor cultuur groei. Side wielen: stappenmotor gecontroleerd schommelen apparaat voor afwisselend onder water en de atmosfeer blootgestelde fase die nodig zijn voor de cultuur groei. (C) Schematische tekening van het coronale rat plakjes gebruikt voor cortex-VTA co-culturen. Cortex secties (links) en middenhersenen secties (midden, rechts) met de VTA ventrale tegmentale gebied (VTA, grijs) worden verkregen door te snijden langs de gebroken lijnen. CTX: cortex; wm: witte stof, cpu: striatum; VTA: Pons: pontine gebied. Zie ook de bijbehorende coronale borden 8, 18, en 20 bij 15. (D) Plaatsing en de groei van een enkel-cortex VTA co-culturen op het MEA en de ontwikkeling over de eerste negen DIV in de cultuur. Let op de afvlakking van de cultuur en de geleidelijke uitbreiding op de array. Reflecterend weefsel onderdelen geven gedegenereerde cellen en weefsels puin. Gezond weefsel is ondoorzichtig en grijs onder transilluminatie met zichtbaar licht.

Figuur 2. Neuronale lawines in corticale organotypische culturen. (A) Overplot van drie periode van spontane activiteit op de array, gescheiden door een aantal seconden. Merk op dat iedere activiteit periode bestaat uit een negatieve LFP doorbuigingen op veel elektroden op de array (elke kleur labels een activiteit periode). (B) negatieve piek tijden van nLFPs van elke elektrode worden geassembleerd in een raster van activiteit. 'Column'-achtige structuren aangegeven periodes van bijna synchrone activiteit. (C) Merk op dat de kolommen die sterk lijken gesynchroniseerd op een tijdschaal bestaan uit meerdere kolommen op de hogere temporele schalen (3 tijdsschalen afgebeeld). (D) Schematische weergave van de neuronale lawine algoritme. Op een 2 x 2 elektrode-array piek tijd en amplitude van negatieve LFP doorbuigingen (nLFP) het oversteken van een drempel van-x SD van het geluid worden geïdentificeerd. Spatiotemporele organisatie van nLFPs is geclusterd in achtereenvolgens actieve tijd bakken van de breedte At. De grootte van een cluster wordt aangeduid met het aantal actieve sites, dat wil zeggen elektroden met nLFP (s = 4), of de geïntegreerde som van nLFP amplitudes (s = 130 uV). De levensduur wordt gemeten in veelvouden van At. (E, F) Power wet in cluster grootte-verdeling identificeert clusters als neuronale lawines. Merk op dat de keuze voor een bepaalde interelectrode afstanden Δd voor de array (hier 200 urn) een bepaald At waarbij de dynamiek moet in acht worden genomen introduceert. Meer specifiek, de verhouding waarin Δd / At benadert de gemiddelde voortplantingssnelheid in het netwerk, waarbij de helling α van de macht wet benadert -1,5 voor de neuronale lawines 2,4,5. Gelieve Klik hier voor een grotere versie van figuur 2 te zien.
Discussion
1. Technische kwesties:
- Steriele techniek. De voorbereiding van de MEA's en de cultuur voorbereiding zijn allemaal uitgevoerd in een laminaire stroming kap onder steriele omstandigheden. Antibiotica, die neuronale activiteit beïnvloeden, worden niet gebruikt op elk moment tijdens de voorbereiding en de cultuur proces.
- Plasma / trombine coagulatie en weefsel hechting op de MEA. Tissue overleven op de MEA vraagt om een zorgvuldige balans tussen de tijd die nodig is voor plasma / trombine stolling en de tijd van weefsel blootstelling aan de atmosfeer. Een korte stollingstijd risico's vroegtijdig loslaten van delen van de MEA, terwijl langdurige blootstelling aan de atmosfeer triggers weefsel degeneratie. Omdat de kracht van de trombine-oplossing bepaalt de snelheid van de coagulatie-proces, het is een zeer belangrijke parameter voor het succesvol bevestigen, gezonde culturen aan de MEA oppervlak. We krijgen de beste resultaten met behulp van 1000 eenheden (1KU, een NIH-eenheid = 0,324 ± 0,073 mg). Belangrijk is, onvolledige mengen van de plasma / trombine-oplossing resulteert in een ruimtelijk heterogene stolling bevorderen van plasma-breuken tijdens het kweken. Deze plasma 'gaten' ernstig aantasten kunst gezondheid en losmaken van de cultuur van micro-elektroden, waardoor afbreuk te doen aan de kwaliteit van de elektrofysiologische opname. Werken met koelplaten tijdens de MEA / weefsel montage vertraagt de bloedstolling en zorgt voor homogeen mengen van de plasma / trombine-oplossing en een goede positionering van de weefselcoupes.
Op dezelfde manier, door het toevoegen van medium in enkele druppels aan de cultuur kamer om onder te duiken de cultuur na slechts vijf minuten van coagulatie, vermindert de kans op weefsel detachementen te wijten aan de oppervlakte spanningen. Een succesvolle cultuur plat op de MEA en enigszins uit te breiden met een gezonde groei voor week, zonder noemenswaardige tekenen van onvolledige weefsel-MEA contact of weefsel degeneratie (bijv. Fig. 1D). - Tissue dissectie. Micro messen zijn sterk verbeterd onze dissectie proces. We maken gebruik van dubbele rand scheermesjes (Fine Science Tools - breekbare scalpelmesjes - 100050-00), waaruit we splitsen ~ 2 mm breed 'messen' met behulp van een tang, en houd ze met een scalpel houder. Weefselmonsters ontleed van de coronale slice met behulp van een gladde, verticale vaste beweging van het blad daardoor sterk verminderen van de mechanische stress door te trekken van het weefsel algehele verbetering van de cultuur van de gezondheid.
- Tissue koeling. Een goede koeling van de plakken en weefselcoupes tijdens de voorbereiding is essentieel voor de cultuur succes. We maken gebruik van op maat gemaakte koude schotels gebouwd van Peltier-elementen die bevestigd zijn aan onder een metalen schijf. De warmte geproduceerd door de Peltier-element wordt verwijderd door middel van koud water perfusie. Dit vermindert de voorbereidingstijd en standaardiseert koeling tijdens elke fase van de voorbereiding (Dold Labs & Engineering 131 Plantation Dr Seguin, TX 78155;. (830) 560-1.471)).
- Incubator conditie. Een op maat gemaakte incubator met een precieze interne rockende voorwaarden is cruciaal voor ons succes in het cultiveren van plakjes op MMO's. Op basis van een origineel in-house ontwerp van Multichannelsystems (momenteel niet in de handel verkrijgbaar), de interne schommelen apparaat bestaat uit trays die verbonden zijn aan twee grote wielen. Stappenmotoren en geautomatiseerde controles zorgen voor een nauwkeurige rockende traject (schommelen hoek, schommelen snelheid, en intermitterende pauzes). Uiteindelijk, slice culturen moeten worden blootgesteld aan sfeer en cultuur medium in slow afwisseling. De traditionele benadering is om slice culturen plaatsen in smalle buisjes die langzaam draaien langs daar langste as. Hier, is traagdraaiend produceert geen mechanische stress als gevolg van de rotatie zelf, en de rotatie snelheid is hoog genoeg om een optimale 'voeden / ademhaling'-cyclus van ongeveer 5 te verkrijgen - 10 min. duur. De meer compacte interieur van de MEA kamer, het totale volume van ~ 2 cc, en de kleine kweekmedium volume nodig voor medium conditioneren door het weefsel zelf, vormt een belangrijke uitdaging. Door het schommelen van de MEA tussen ± ~ 70 ° hoek (cyclustijd: ~ 200 s), het vertragen van de snelheid schommelen als de cultuur overgangen tussen de vloeibare fase en de atmosfeer, en het stoppen van de schommelen in het uiterste hoeken voor een langere blootstelling aan de atmosfeer heeft essentieel geweest voor de cultuur te overleven.
2. Ontwikkelingsleeftijd van cortex culturen neuronale lawines studeren
Acute segmenten van de rat cortex worden over het algemeen genomen op PND 0-1 en gekweekt voor veel weken op de MEA. Vroege studies hebben duidelijk aangetoond dat enkele cortex slice culturen, na een aantal weken in vitro, een gelaagde structuur met herkenbare celtypen die gemakkelijk kunnen worden vergeleken met in vivo cel klassen 18,18-21 te behouden. De gelaagde organisatie in dit in vitro systeem is handig gebruikt om de thalamus studieinnervatie van cortex tijdens de ontwikkeling 22-24, evenals voor het rijden subcorticale structuren, zoals het striatum 25,26. In feite is de specificiteit in de vorming van neuronale verbindingen binnen en tussen hersengebieden zorgt voor de bouw van complex in vitro systemen die een groot aantal gedetailleerde projectie systemen heroveren, bijvoorbeeld dat van de cortex-basale ganglia circuits 27-30.
Na 4 - 6 weken in vitro, single cortex plakjes 31 en cortex plakjes samen gekweekt met striatum 26 of 32 thalamus vertonen spontaan omhoog en omlaag staten meestal gevonden in vivo in het urethaan verdoofde rat 33. De fijne temporale organisatie van deze up-staten draagt het stempel van geneste θ-en γ-oscillaties indicatief voor een elektrofysiologische volwassen netwerk van piramidale neuronen en fast-additie GABAergische interneuronen 31. Belangrijk is dat bij het ontbreken van dopamine D2-receptor stimulatie, is de rijping van parvalbumin-positieve corticale interneuronen vertraagd met ongeveer 2 weken in cortex slice culturen 34. In lijn met deze bevindingen, de ontwikkeling tijdsverloop van geneste θ-, β-en γ-oscillaties komt overeen met in vivo wanneer cortex schijfjes zijn co-gekweekt met het ventrale tegmentale gebied (VTA), die dopaminerge neuronen projecteren naar het bevat cortex 6.
Deze studies geven aan dat bij het bestuderen van neuronale lawines, die hoge mate afhangen van volwassen snel GABA-erge inhibitie en bevinden zich in de oppervlakkige lagen van de cortex 2,4, grote zorg moet worden genomen om een goede rijping van de corticale weefsel te verzekeren. Terwijl de neuronale lawines ontstaan in enkele cortex culturen over het tijdsverloop van 2 - 5 weken 4, wanneer die een ontwikkelingstijd cursus die is afgestemd op de in vivo ontwikkeling, cortex plakjes behoefte aan een passend dopamine-receptor stimulatie, bijvoorbeeld door co-kweken cortex plakjes met de VTA 6.
Disclosures
Geen belangenconflicten verklaard.
Acknowledgments
Deze studie werd gefinancierd door de afdeling van de intramurale Research Program (DIRP) van het National Institute of Mental Health, National Institutes of Health.
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}