

Summary
대뇌 피질의 임계 상태 역학 지표의 연결 눈사태, 즉 스케일 - 불변량 spatio - 관자놀이 활동 파열을 연구하는 강력한 방법입니다. 눈사태 정확하게 제어 조건 하에서 평면 통합 다중 전극 어레이 (MEA)와 활동의 장기 측정을 허용 교양 피질의 표면 레이어를 개발에 자발적으로 등장.
Abstract
대뇌 피질도 특정 입력 또는 모터 출력의 부재에 저절로 활성화됩니다. 개발하는 동안,이 활동은 마이 그 레이션과 차별화 피질 세포 유형의 연결과 연결 1의 형성에 중요합니다. 성숙한 동물에서 지속적인 활동이 과거와 감각 자극이 완벽하게 미래의 행동을 계산하기 위해 통합되는에 동물의 현재 상태를 반영합니다. 따라서, 지속적인 즉, 자발적인 활동의 조직의 명확한 이해 피질의 기능을 이해하는 필수 조건이다.
수많은 레코딩 기술은 피질에 지속적인 활동이 그의 개인적인 활동 transiently 세포 microelectrodes와 지역 현장 잠재력 (LFP)에서 감지가 가능하다 더 큰 이벤트 합계, 또는 뇌파 (EEG), magnetoencephalogram (MEG 많은 뉴런으로 구성됩니다 것으로 나타났다 ), 그리고 기능적인 자기 공명 영상 (fMRI)에서 대담한 신호. mesoscopic 규모에서 높은 시간적 및 공간적 해상도 (뉴런의 여러 수천)으로의 연결 인구 활동을 공부하면 LFP은 현재 선택의 방법입니다. 세포외 microelectrode에서 로컬 microvolts의 수백까지 LFP 급격한 deflections의 공간 neighbored 뉴런 결과의 활동을 동기화. microelectrodes의 배열을 사용하는 경우, 이러한 deflections의 단체가 편리하게 공간과 시간에 모니터링할 수 있습니다.
의 연결 눈사태는 뇌 2,3에 지속의 연결 활동의 규모 - 불변량 spatiotemporal 조직을 설명합니다. 그들은 anesthetized 쥐 6 생체내에서 체외 4,5 년 설립 피질의 표면 레이어에 특정되며, 정신 원숭이 7인치 중요한 이론적 경험적 연구 모두 2,8-10는 눈사태의 연결 정보 전송 및 정보 처리를 최적화 피질의 정교하게 균형 임계 상태 역학을 나타내는 것이 좋습니다.
그들이 정확하게 제어 조건 하에서 눈사태 활동의 안정적인 레코딩을 허용으로의 연결을 눈사태 개발, 유지 보수 및 규제의 메커니즘을 연구하기 위해 체외 준비에 매우 유용합니다. 현재 프로토콜은 평면 통합 microelectrode 배열 (;도 11-14을 참조하십시오 MEA)에 성장 organotypic 피질의 문화, 즉 슬라이스 문화에서 표면 레이어 개발을 활용하여 체외로의 연결을 눈사태을 공부하는 방법에 대해 설명합니다.
Protocol
1. 장기 레코딩을위한 MEA와 살균, Sealable 유리 회의소
- 안전하고 꽉 문화 챔버 폐쇄에 필요한 테플론 - 플라스틱 캡 (에이스 유리)와 스레드 유리 실린더는 스레드 (그림 1A, B)의 하단에서 (Aceglass) 약 2mm를 잘라있다. 물 (3 배) 200 증명 에틸 알콜에 5 분 끓는과 rinsing하여 깨끗한 유리 반지가 건조하게.
- 나누어지는 실리콘 솔루션은 MEA의 표면에 유리 반지를 첨부해야합니다. Sylgard 184 실리콘 엘라스토머 키트의 A & B는 철저하게, -20 ° C. 1 ML의 aliquots에 저장, 공기 방울을 제거 15 분 앉아 보자 부품의 15 ML를 섞어
- MEA에 접착제 유리 반지 (8x8 그리드 W / 내부 접지 전극, 쥐 / 마우스 30 μm의 전극 직경 100분의 200 μm의 전극 간의 거리) (그림 1A, B). 작은 게이지 바늘로 주사에 실리콘 1 ML (23 ° C)를 차지. ~ 60 ° C 뜨거운 접시에 2 시간 - 유리 반지, MEA의 중심 유리 반지, 강한 인장을위한 반지의 외부 주위에 실리콘의 추가 계층을 적용할의 unpolished 컷 표면에 실리콘을 적용, 1 치료하자.
- 배 70 % 알코올에 의해 다음 탈이온수에 씻어하여 층류 후드의 MEA 챔버와 챔버 뚜껑을 소독 (3 X, 지난 알코올에 10 분 앉아 보자 린스) 자외선 10 챔버 분 노출과 캡 내부 다음 . 압력솥 MEA 챔버 (120 ° C, 서부 유럽 표준시, 45 분)하고 건조하게.
- 폴리 - D - 라이신과 코트 MEA 표면 문화 챔버 내부. 들어 오히려 lipophilic하는 새로운 MEAs, 전극 그리드에서 솔루션의 반복 비말의 열망에 의해 외투. 사용 MEAs 들어, 솔루션과 챔버 바닥 커버, 여분의 액체를 대기음, 판상 후드 안쪽 무균 조건 하에서 증발 수 있습니다. 저장 및 향후 사용을위한 MEA 챔버를 밀봉하기 위해 뚜껑을 첨부합니다.
2. Organotypic 문화의 준비 및 성장에 필요한 재료
- 무균 배양 접시 (팔콘, 100 x15; ~ 5mm 수준)에 부어, 0.9 % NaCl에 살균 한천을 디졸브, 식지, 스토리지에 대한 Parafilm과 멸균 포장. 사용하기 위해 단단한 한천에서 20 X 10 X 5mm 블록을 잘라.
- 포장 스토어 슈퍼 접착제, 예를 들어 Devcon 슈퍼 접착제 II는 불임을 보존하기 위해 층류 후드 안쪽 열기 전에 70 % EtOH로 닦여.
- 드 - 이온화된 문화 물 (시그마) 40 ML에 포도당의 40g을 추가 50 % d 개의 포도당 (시그마 초소형, G7528)를 준비합니다. -20 ° C. 2 ML aliquots에 저장
- Gey의 밸런스드 소금 솔루션 500 ML 50 % D - 글루코오스 4 ML을 추가하고 사용하기 전에 냉동실에 윤활제 (액체 / 얼음 결정의 혼합물)에 진정해.
- 부드럽게 10 분, 소용돌이 및 멸균 페트리 접시에 명확한 내용을 decantate - 5 ML 드 이온화 문화 물 (거품 형성을 방지, 부드러운 악수)에서 Reconstitute 치킨 플라즈마는 솔루션 5 쉬라 구. 무균 필터 (0.22 μm의 기공 필터, 단백질) -20 ° C.에서 플라즈마 솔루션 cryotubes로 나누어지는 350 μl (NuncTM), 저장
- (NuncTM) 나누어지는 40 μl cryotubes로 (0.22 μm의 기공 필터) 따라서, 살균 필터 소 플라즈마에서 Reconstitute의 트롬빈, -20 ° C.에 저장 솔루션을 작업하기 위해 Gey의 균형 소금 솔루션 W / D - 포도당의 375 μl의 트롬빈 용액 40 μl를 희석.
- 말 혈청, 200 ML 기초 중간 이글, 50 % 포도당 200 MM L - 글루타민 2 ML 4 ML이 추가 100 ML 행크의 버퍼 생리 식염수 100 ML을 혼합하여 배지 400 ML를 준비합니다. 4 8주 100 ML PYREX 병에 ° C. - 4 저장할 수 있습니다
- 0.3 MM의 유리딘, 0.3 MM ARA - C 사이 토신 - β - D - arabinofuranoside 및 0.3 MM 5 - 플루오로 - 2' - deoxyuridine, 살균 필터, 나누어지는 200 μl와 6 -20 ° C에서 저장을 혼합하여 유사 분열 억제제 준비 - 12개월.
3. 접속 및 복부 Tegmental 면적 (VTA) 조직 해부 (시간 : <1 시간)
- 절차 산출 피질과 ~ 12 쥐 또는 마우스의 공동 문화, 그리고 무균 조건 하에서 층류 후드 내부의 준비를 위해 VTA 조직 섹션. 조직 컬렉션 총 시간은 1 시간 이상해야합니다.
- 1 - 2 출생 후의 일 (쿨러), 건강하고, 잘 nourished 새끼를 (복부 '우유 명소'의 존재를 쓰레기 사이즈 ~ 10) 가져가라. 주둥이로 부드럽게 강아지를 기다려, 그것이 자유롭게 끊지 수 있도록하고, 신속하게 날카로운 가위로 목 기지에서 목을 베다.
- 뇌 제거 들어, (두 측면 가위 인하) 피부를 제거하는 미세 안과 가위 (1 sagital 정중선 컷, 1 코로나 피질에서 절단 / 소뇌 접합)와 오픈 두개골을 잘라. 모든 4 해골의 날개를 다시 뒤집기. 날카롭게 주걱으로 caudally 두뇌 아래, 후각 망울을 통해 frontally 사전 주걱 잘라. 부드럽게 두개골의 두뇌를 리프트하고 빠른 냉각 및 임시 저장 살균, 냉각, Gey의 솔루션으로 보내드리겠습니다. 2 개 두뇌를 위해 3.3 단계 3.2 반복 (총 시간 : <20 분).
- VTA를 얻으려면조직, 작은 주걱을 사용하여 살균, 건조 페트리 접시에 두뇌를 전송합니다. 더 부드럽게 대해 1cm 옆으로 각각의 머리를 슬라이딩하여 여분 액체를 제거합니다. 면도날을 사용하여 소뇌의 수준에서 코로나, 수직 절단에 의해 뇌의 줄기를 제거합니다.
- 절차를 깔끔히하는 동안 두뇌의 기계적 안정화를위한 디스크를 장착에 대한 글루 한천 블록. superglue의 얇은 라인을 플레이스 디스크에 한천 블록의 앞에 몇 밀리미터 (한천을 감동 접착제를 피하기).
- 작은 주걱을 사용하여, 아래, 정면 기둥을 전송하고 각 두뇌를 탑재합니다. 정면 기둥이 설치 디스크에 접착하는과 복부 양쪽이 도중 컷 조각의 절단하고 쉽게 이륙. 적절한 기계적 안정화를 달성하기 위해 어떤 접착제 잔류물없이 한천을 만지지 있는지 확인
- 조심스럽게 vibratome 트레이에 뇌 어셈블리 (예 : Leica VT1000)과 잠수함 및 보안 장착 디스크 살균, 냉각 Gey의 솔루션으로 가득. 조심스럽게 청소 면도날 (90% EtOH)으로 가장 높은 진동 주파수에서 midbrain의 코로나 조각 잘라 400 500 μm의의 두께에 상대적으로 낮은 앞으로 속도. 흡입 전구로 바뀌 파스퇴르 피펫을 사용하여 전송 및 살균, 냉각 Gey의 솔루션으로 가득 35 X 10mm 배양 접시에 VTA 포함하는 슬라이스 수집 (그림 1C을, 또한 코로나 판 18 참조 - 15 E22 20)를.
- 에 대한 대뇌 피질 섹션 단계를 반복합니다 3.2-3.6지만, 피질과 소뇌 사이에 수직 절개를 적용하고, 정면 기둥과 forebrains을 탑재합니다. striatum의 수준에서 시작 약 3 코로나 조각 (350 μm의 두께)가 향후 피질의 절개를 위해 수집됩니다.
- 부러진 면도날 만든 마이크로 칼을 사용하여 전두엽 피질과 stereomicroscope 아래 VTA (그림 1C)이있는 midbrain 영역의 ~ 2mm 폭 코로나 섹션을 해부하다. 냉장 Gey의 솔루션으로 가득 작은 접시 (예 : 챔버 슬라이드)에서 별도로 조직 섹션을 수집합니다.
4. MEA에 접속하고 VTA 조직 조각 장착 (시간 : <1 시간)
- 초점에있는 전극 배열을 가진 스테레오 현미경으로 실온에서 위치 MEA합니다. 센터 25 깨끗하고 먼지 무료 플라즈마의 μl 비말 및 살균 전극 배열 매트릭스. 작은 spatulae를 사용하면, 조심스럽게 플라즈마 비말에 피질과 VTA 부분을 슬라이드.
- 냉각 판에 MEA를 삽입,보기를 집중할 플라즈마 비말에 트롬빈의 25 μl를 추가 후, ~ 15 s에 대한 진정하자. 트롬빈 피펫 팁을 사용하여 조심스럽게 MEA에 걸쳐 작은 원형의 움직임과 플라즈마 / 트롬빈 혼합물을 확산. 직접 취성 전극 어레이를 만지지 마십시오. 부드럽게 배열의 두 번째 전극 행에 따라 지느러미 테두리와 배열에있는 피질을 위치. 이 방법의 개발 표면 레이어는 결국 배열의 알림을 커버합니다. VTA는 피질 부분의 복부 국경 (그림의 1D)에 인접 배치됩니다.
- 뚜껑과 느슨하게 MEA / 문화 조립 플라즈마 / 트롬빈 응고 수 있도록 실내 온도에서 후드 안쪽 ~ 5 분 동안 앉아 높은 습도를 유지하는 MEA 챔버를 닫습니다. 3 더 많은 문화 4.3 - 한편, 단계 4.1를 반복합니다.
- 조심스럽게 25 X 8분의 5 바늘로 1 CC의 주사기를 사용하여 문화 챔버에 작은 물방울의 문화 매체 600 μl를 추가합니다.
- 단단히 MEA 챔버 장소 MEA / 보육 (그림의 1B) 내부의 락을 보관 트레이에 문화 어셈블리를 닫습니다. 하려면, 3의 절차를 빠르게 - 4 MEAs는 중복 시퀀스에 조립하실 수 있습니다. 12 MEAs에 대한 어셈블리 시간 <1 시간해야합니다.
- 관내 (DIV) 2 일 후에, 유사 분열 억제제의 10 μl를 추가합니다. 새로고침 문화 미디어 네 DIV 60 % 및 이후 매 4 일까지.
5. Electrophysiological 녹화 및 자극 생성
- MEA의 각 전극에서 ~ 10 분 24 kHz에서시 ~ 일주 5,6 기록 자발적인 활동 후, 지역 현장 잠재력 (LFP)와 행동 잠재력을 해고 뉴런의 경향에 상당한 deflections 사이의 관계를 확립하려면 (하드웨어 : MEA1060 W / 블랭킹 회로, x1200 이득, 12 비트 A / D, 범위 0-4096 뮤직 비디오, 멀티채널 시스템, 소프트웨어 : MC_Rack). 접지는 내부 접지 전극 통해서 제공, 또는 외부 자세 / AgCl 반 세포를 추가하여.
- 세포외 스파이크 활동 200 Hz에서 (밴드 패스 300 - - 3,000 Hz에서) 1의 밴드 패스 필터 LFP 구분합니다. 스파이크 활동이 더욱 오프라인 스파이크 sorters (예 : Plexon 주식 회사)를 사용하여 단일 및 다중 유닛 활동으로 나눌 수있다. 계산 스파이크는 각 전극에 대한 평균을 촉발시켰다. 대뇌 피질의 문화, 대부분의 평균은 문화의 연결 급상승의 선호하는 시간을 제외 LFP의 deflections (nLFP)를 식별합니다.
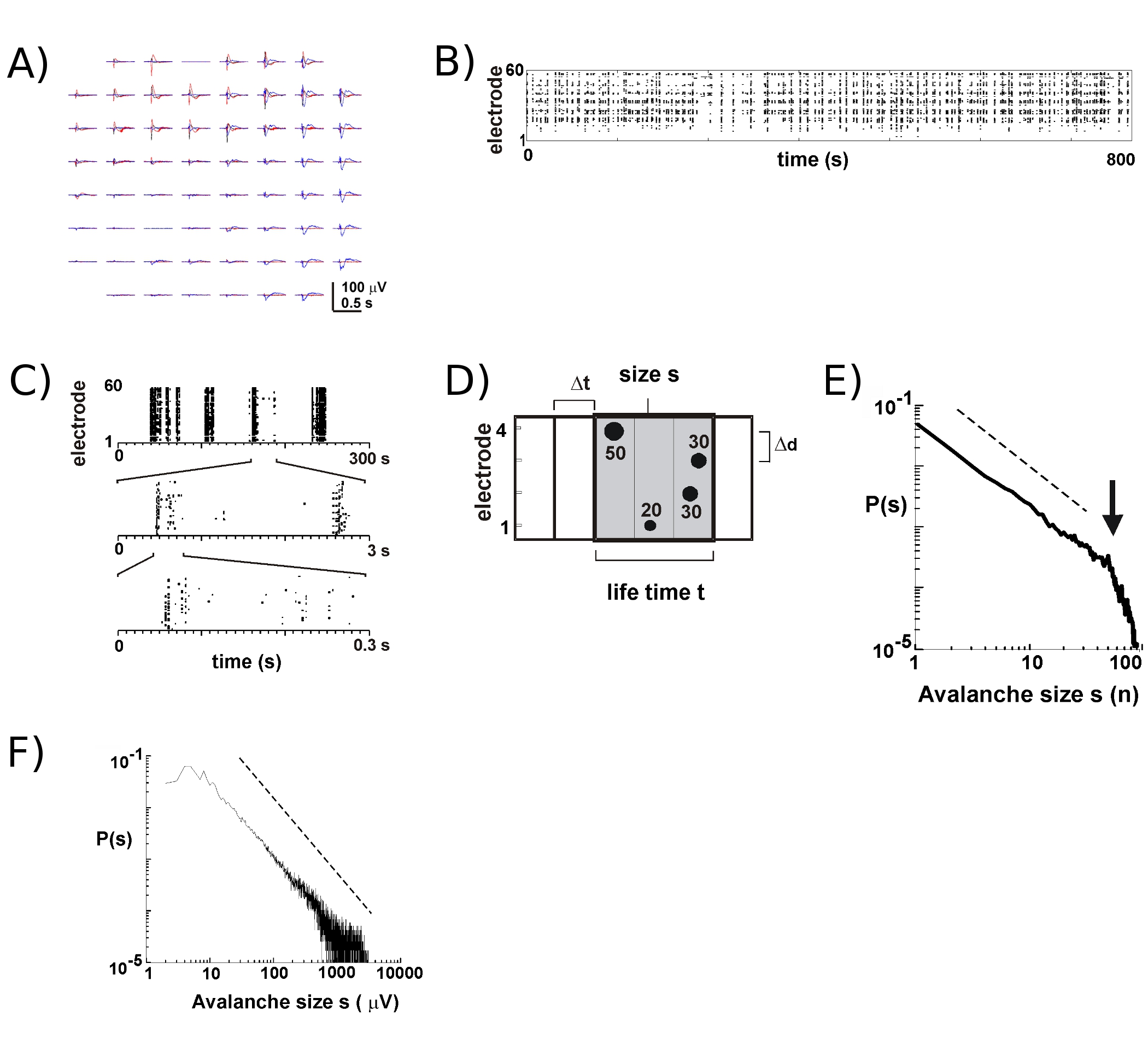
- (그림 2) 각 전극에 대한 LFP 추적에서 소음 (SD)의 -3 표준 편차의 임계값을 계산, nLFPs의 최대 시간과 amplitudes을 결정 십자가 임계값 (그림 2B, C). 세부 2,4 참조를 위해, 그리고 길이 Δt의 연속 시간 빈스 (그림 2D 같은에있는 모든 전극에서 nLFPs을 합치하여 배열에 spatiotemporal nLFP 클러스터를 식별 - 시간 빈 Δt를 (8 MS을 2 사이 등) 선택 , 5).
- 눈사태의 연결을 식별하기 위해 각 nLFP 클러스터의 크기를 계산, 활성 전극 또는 nLFP amplitudes의 합계의 예 : 숫자, 크기 히스토그램을 구축하고, 이중 로그 좌표 플롯. 의 연결 눈사태 들어, 크기 분포 이중 로그 좌표 2 (그림의 2E, F)의 직선에 의해 approximated 전원 법을 따른다. 전원 법에 대한 통계 시험 16를 참조하십시오.
- 진폭 S와 전류 제어 자극이 (자극 발생기 STG 1008, 멀티채널 시스템)이 적용되는 전극을 통해를 선택하여 조직에서 evoked 반응을 이끌어내는. - 200 μA 진폭 - S와 50 μs 10 사이의 진폭 + S / 2 S 100 μs 다음 : 전극의 손상을 줄이기 위해 제한된 범위 바이폴라 사각형 파형 단일 충격을 담당하고 중립적인 자극을 사용합니다. 자세한 내용은 설명서를 참조하십시오.
- 동적 범위 9 자극 후 500 MS를 따라 모든 전극 4 kHz에서 샘플링 속도에서 LFP 응답을 기록 기록 자극을 기록합니다. 자극 유물을 절감하고 사전 증폭기 포화를 방지하기 위해 자극하는 동안 머리를 무대 앰프를 연결을 끊으 회로 (멀티 채널 시스템) 블랭킹 사용합니다.
6. 대표 결과 :
8 관한 새로운 MEAs와 함께 - 9 밖으로 10 문화의 여러 주 동안 살아남을 것입니다. 장기 녹음의 대부분은 우리가 많은 주 과정 5 이상의 개별 문화의 발전을 따라 수 있도록 문화 매체에 배양기 안에서 이루어집니다. 우리의 실험을 바탕으로, LFP 레코딩이 안정적으로 100 개 이상의 문화 일간 사용 MEAs와 함께 얻을 수 있습니다. 대조적으로, 세포 스파이크 활동보다 안정적으로 비교적 새로운 MEAs (<40 문화 일)와 함께 측정됩니다. 전형적인 실험에서는, 우리는 문화 챔버가 봉인 유지 붙어있는 머리 단계 (그림 1B, 왼쪽)와 트레이에 저장 트레이 (그림의 1B, 오른쪽)에서 MEA를 전송합니다. 피질 5, 피질 - VTA 공동 문화 6,뿐만 아니라 anesthetized 쥐 6과 생체내 7 깨어 원숭이, 표면 레이어로의 연결을 눈사태 동안의 연결을 해고에서와 같이 주로 LFP의 정상 부정적인 편향에 가까운 발생 (nLFP ). 따라서, 로컬 동기화의 연결 그룹의 spatiotemporal 조직 배열 17 공간과 시간에 nLFPs의 발생을 측정하여 예상하실 수 있습니다.
MEA의 활동 일시적 클러스터에 등장하는 경향이 하나의 전극에서 그러한 활동을 다른 사이트에서 활동을 함께합니다. 이러한 활동 기간 동안 LFP의 전형적인 파형은 몇 초 간격을 발생 3 클러스터를 계획하여 그림 2A에 표시됩니다. 각 클러스터에 대한, 부정적인 현장 deflections은 1 창 안에 여러 개의 전극에서 볼 수 있습니다 S. 여러 부정적인 SD의 문턱을 건너 nLFP의 봉우리를 추출하면, nLFP 피크 시간의 형태로 활동이 점들의 '열'가 다양한 전극 (그림 2B)에 일치하는 nLFPs 근처 대표하는 래스터에서 편리하게 시각이다. 이 활동의 spatiotemporal 조직은 다소 복잡하다, 더 많거나 적은 균일한 낮은 시간적 해상도에 나타나는 '열'는, 높은 해상도와 세속 등등 (그림 2C)에서 별도의 클러스터로 구성되어 있습니다. 사실, spatiotemporal nLFP 클러스터의 출현은 매우 대뇌 피질의 네트워크로 구성됩니다. 보다 구체적으로, 조직의 연결 눈사태를위한 스케일 - 불변량입니다. 이것은 주어진 시간적 해상도 Δt에서 클러스터 크기의 확률을 계산하여 입증됩니다. 여기서 클러스터는 동일하거나 연속된 시간 용기 (그림 2D)에서 발생하는 nLFPs 구성되어 있습니다. 이러한 클러스터의 크기가 클러스터 당 nLFPs 또는 클러스터 당 통합 nLFP amplitudes의 총수로 표현되면, 클러스터 크기 배포판은 누구 기울기 -1.5 2,4,5,7로 표시되었습니다 전원 법을 공개 ( 그림. 2E, F). 이 배포판은 사이즈 S K는 상수 요소이다 kxs에의 비율입니다 클러스터 크기의 규모 - 불변량 주문을 식별하는 것을 참고, S.의 독립 -1.5 K입니다 이 전원 법 조직이 배열 2 크기의 독립, 시간적 해상도 Δt 2, 상당한 nLFP를 식별하는 데 사용되는 임계값 7 deflections. 의 연결 그룹 크기 7 nLFP 진폭의 비늘은 nLFPs의 규모 - 불변량 조직 규모 - 불변량, 즉 프랙탈, 주문 O를 반영하기 때문에F 로컬 모든 크기를 포함의 연결 그룹을 동기화.

그림 1. (A) 스레드 유리 반지와 MEA의 측면과 상단 전망 탑재하고, 해당 모자. (B) 보육의 전망 인사이드. 왼쪽 headstage는 인큐베이터 조건에 따라 하나의 문화에서 녹음 수 있도록 탑재합니다. 오른쪽 : 문화의 성장을 위해 수많은 MEAs을 잡고 트레이. 사이드 바퀴 : 문화의 성장에 필요한 잠수 분위기 노출 단계를 교체를위한 모터 제어 락을 장치를 스테핑. 대뇌 피질 - VTA 공동 문화에 사용되는 코로나 쥐 조각을위한 (C) 도식 도면. VTA 복부 tegmental 지역 (vta, 회색)이있는 컴퓨터에 접속 섹션 (왼쪽)와 midbrain 섹션 (중간, 오른쪽)은 깨진 라인을 따라 커팅에 의해 얻을 수 있습니다. CTX : 대뇌 피질, WM : 흰색 문제, CPU : striatum; vta : 폰스 : pontine 공간이 있습니다. 15도 해당 코로나 접시 8, 18, 20를 참조하십시오. (D) 위치 및 MEA와 문화의 첫 번째 9 DIV를 통해 자사의 개발에 단일 피질 - VTA의 공동 문화의 성장. 문화와 배열에 미치는 진보적인 확장의 평평화을 확인합니다. 반사 조직 부품 퇴화한 세포 조직 파편을 나타냅니다. 건강한 조직은 가시 광선과 transillumination 아래의 불투명과 회색빛이 있습니다.

그림 2. 피질 organotypic 문화에의 연결을 눈사태. (A) 배열에 대한 자발적인 활동의 세 시대의 Overplot는 몇 초으로 구분합니다. 각 활동 기간은 배열에 많은 전극에 부정적인 LFP의 deflections (각 색상 하나의 활동 기간을 라벨)로 구성되어 있습니다. (B) 각 전극에서 nLFPs의 부정적 피크 시간은 활동의 래스터로 조립합니다. 'Column' 같은 구조는 근처 동기 활동 기간을 지적했다. (C) 고도로 한 번에 규모에 동기 나타나는 열이 높은 시간적 비늘 (3 시간적 비늘이 그림 참조)에 여러 개의 컬럼으로 구성됩니다합니다. 의 연결을 눈사태 알고리즘 (D) 도식 표현. 소음 SD - X의 문턱을 넘어 부정적인 LFP의 deflections (nLFP)의 2 X 2 전극 배열 피크 시간 및 진폭에 식별됩니다. nLFPs의 Spatiotemporal 조직 폭 Δt의 연속 활성 시간 방식으로 클러스터입니다. 클러스터의 크기가 nLFP과 적극적인 사이트의 수를 즉, 전극에 의해 식별됩니다 (에스이 = 4), 또는 nLFP amplitudes의 통합 SUM (S = 130 μV). 생활 시간은 Δt의 배수 단위로 측정됩니다. 클러스터 크기 배포 (E, F) 전원 법률의 연결 눈사태로 클러스터를 식별합니다. 배열 (여기서는 200 μm의)에 대한 Δd 특정 interelectrode 거리의 선택은 역학을 관찰해야되는 특정 Δt를 소개합니다. 보다 구체적으로, 비율이있는 Δd / Δt는 전원 율법의 슬로프 α는 눈사태의 연결을 2,4,5에 대한 -1.5 대략있는에, 네트워크의 평균 전파 속도를 대략. 하시기 바랍니다 여기를 클릭 그림 2의 더 큰 버전을 볼 수 있습니다.
Discussion
1. 기술적 문제 :
- 멸균 기술은. MEAs 문화 준비의 준비는 모두 무균 조건 하에서 층류 후드 안에서 수행됩니다. 의 연결 활동에 영향을 미칠 항생제는, 준비 및 문화 과정 언제든지 사용되지 않습니다.
- 플라즈마 / 트롬빈 응고 및 MEA의 MEA. 조직 생존에 조직 준수는 플라즈마 / 트롬빈 응고와 분위기 조직 노출 시간에 필요한 시간 사이에 신중 균형이 필요합니다. 분위기를 장기간 노출이 조직 변성을 실행하는 동안 짧은 응고 시간은 MEA에서 섹션의 조기 부대를 위험. 트롬빈 솔루션의 강도가 응집 과정의 속도를 결정하기 때문에, 그것은 성공적으로 MEA 표면에, 건강한 문화를 부착을위한 매우 중요한 매개 변수입니다. 우리는 1000 단위 (, 1 NIH 단위 = 0.324 ± 0.073 MG 1KU)를 사용하여 최상의 결과를 얻습니다. 중요한 것은, culturing 동안 플라즈마 - 파열을 증진 공간 이기종 응고에서 혈장 / 트롬빈 솔루션 결과의 혼합 불완전. 이러한 플라즈마 '구멍'은 심각하게 문화의 건강 영향을 microelectrodes에서 문화를 분리,이를 electrophysiological 녹음의 품질을 저하. MEA / 조직 조립하는 동안 냉각 플레이트로 작업하면 응고를 느려지고 플라즈마 / 트롬빈 솔루션 및 조직 섹션의 적절한 위치의 균일한 혼합을 허용합니다.
마찬가지로, 응고 불과 5 분 후에 잠수함의 문화를 문화 챔버에 하나의 물방울에 미디어를 추가하여 크게 표면 긴장으로 인해 조직 detachments의 위험을 줄여줍니다. 성공적인 문화는 MEA에 평평하고 약간 불완전한 조직 MEA 연락처 또는 조직 변성 (예 : 그림. 1D)의 주요 증상없이 몇 주 동안 건전한 성장을 확장됩니다. - 조직 해부. 마이크로 칼 크게 우리의 해부 과정을 개선했습니다. 우리가 폭 ~ 2mm '블레이드'는 펜치를 사용하여 분리하고, 메스 홀더와 함께 그들을 보유하고있는 경우에서 우리는 더블 에지 면도날을 (100050-00 - - 깨뜨릴 수있는 메스 블레이드 파인 과학 도구)을 사용합니다. 블레이드의 부드러운, 수직 회사 모션을 사용하여 코로나 슬라이스의 해부 조직 샘플은이를 크게 인해 문화 건강을 개선하는 전체 조직의 철수에 기계적 스트레스를 감소.
- 조직 냉각. 준비하는 동안 조각과 조직 섹션의 적절한 냉각 문화 성공을 위해 필수적입니다. 우리는 금속 디스크에 아래 첨부 펠티어 요소의 기본 사용자 정의 - 만든 차가운 접시를 사용합니다. 펠티어 요소에 의해 만들어진 열기는 차가운 물 세례를 통해 제거됩니다. 이것은 크게 준비 시간을 단축하고 준비의 각 단계 동안 냉각 standardizes (Dold 연구소 및 엔지니어링 131 플랜타티온 박사 세귄, TX 78155;. (830) 560-1471)).
- 인큐베이터 상태. 정밀한 내부 로킹 조건 맞춤 제작 인큐베이터는 MEAs의 조각 재배에 성공의 열쇠되었습니다. (현재 없습니다 상용) Multichannelsystems하여 자체 디자인 원본에 따라, 내부 로킹 장치는 두 개의 큰 측면 바퀴에 연결된 트레이로 구성되어 있습니다. 스테핑 모터와 컴퓨터 제어 (각도를 락을 속도를 락을하고, 간헐이 일시 중지) 정확한 로킹 궤도에 대한 수 있습니다. 결국, 슬라이스 문화 느린 교대의 분위기와 문화 매체에 노출해야합니다. 전통적인 접근 방식은 천천히가 긴 축을 따라 회전 좁은 튜브로 슬라이스 문화를 배치하는 것입니다. 여기 천천히 회전은 회전 자체에 의한 기계적 스트레스를 생산하지 않으며, 회전 속도는 약 5 최적의 '수유 / 호흡'사이클 얻을 정도로 높은 - 10 분 시간을. MEA 회의소 ~ 2 CC 자사의 총 부피, 그리고 조직 자체에 의해 중간 조절에 필요한 작은 문화 매체 볼륨의 더 많은 콤팩트 인테리어는 상당한 도전 포즈. ± ~ 70 ° 각도 (사이클 타임 : ~ 200 S) 사이의 MEA 로킹으로, 액상과 분위기를 사이에 문화의 전환으로 락을 속도가 느려지고, 그리고 분위기 확장 노출에 대한 극단적인 각도에서 흔들어 중단하는 것이있다 문화 생존을위한 필수되었습니다.
2. 의 연결 눈사태를 공부 피질 문화의 발달 연령
쥐의 대뇌 피질에서 급성 조각은 일반적으로 쿨러 0 가져옵니다 - 1 MEA에 많은 주 동안 교양. 초기 연구는 명확하게 하나의 피질 슬라이스 문화는 체외에서 몇 주 후에 쉽게 생체내 세포 클래스 18,18-21에 비교할 수있는 식별 세포 유형 계층 구조를 유지하는 것을 증명하고있다. 체외 시스템이 계층 조직은 편리 thalamic을 연구하는 데 사용되었습니다개발하는 동안 22-24 피질의 innervation뿐만 아니라 같은 striatum 25,26으로 subcortical 구조를 운전. 사실, 뇌 영역 이내에 전체의 연결 관계 형성의 특이성은 예를 들어 그 피질 - 기저 신경 회로 27-30의 다양한 세부 프로젝션 시스템을 느껴볼 체외 시스템에 복합 건설을 허용합니다.
. 체외, 단일 피질의 조각 31 피질의 조각 striatum 26 또는 시상 자발적인 32 쇼와 함께 공동 교양 위 아래로 일반적으로 우레탄 anesthetized 쥐 33 생체내에서 발견 주에서 6 주 - 4 이후 이러한 최대 - 미국의 벌금 시간적 조직의 특징을지지 중첩된 θ 및 γ - oscillations 피라미드 뉴런과 빠른 속도로 급상승 GABAergic interneurons 31 electrophysiologically 성숙 네트워크의 암시. 중요한 것은, 도파민 D2 - 수용체 자극의 부재에서 parvalbumin 양성 피질 interneurons의 성숙은 피질의 슬라이스 문화 34 약 2 주간에 의해 지연될 수 있습니다. 이러한 결과의 발달 시간 코스 라인에 중첩 θ -, β 및 피질의 조각이에 돌출 dopaminergic 신경이 들어있는 복부 tegmental 지역 (VTA)와 공동으로 양식하는 경우 γ - oscillations는 생체내에 일치합니다 피질 6.
이러한 연구는 결정적인 성숙 빠른 GABAergic 억제에 따라, 피질 2,4의 표면 레이어에있는 눈사태의 연결을 공부하면 아주 신경은 대뇌 피질의 조직의 적절한 성숙을 보장하기 위해 이동하는 것을 나타냅니다. 의 연결 눈사태가 2 시간 코스 이상의 단일 피질 문화에서 발생하는 동안 - 생체내 개발에 일치하는 발달 시간 과정을 필요로 할 때 오주 4, 피질의 조각이 공동 culturing의 피질의 조각으로 적절한 도파민 수용체 자극, 예를 들어 필요 VTA 6.
Disclosures
관심 없음 충돌 선언하지 않습니다.
Acknowledgments
이 연구는 정신 건강의 국립 연구소, 건강의 국립 연구소의 교내 연구 프로그램의과 (DIRP)에 의해 재정 지원되었다.
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}