

Summary
Un metodo efficace per studiare valanghe neuronale, cioè scala-invarianti spazio-temporale scoppia attività, indicativa delle dinamiche condizioni critiche nella corteccia. Valanghe emergere spontaneamente nello sviluppo di strati superficiali della corteccia cerebrale in coltura che permette di misure a lungo termine delle attività con gli array planare integrata multi-elettrodi (MEA), a condizioni ben controllate.
Abstract
La corteccia è attiva spontaneamente, anche in assenza dei fattori di produzione o di uscita del motore. Durante lo sviluppo, questa attività è importante per la migrazione e la differenziazione dei tipi di cellule della corteccia e la formazione di connessioni neuronali 1. Nel animale adulto, l'attività in corso riflette il passato e il presente stato di un animale in cui gli stimoli sensoriali sono perfettamente integrati per calcolare le azioni future. Così, una chiara comprensione dell'organizzazione del corso dell'attività ovvero spontanea è un prerequisito per capire la funzione della corteccia.
Tecniche di registrazione ha rivelato che numerose attività in corso nella corteccia si compone di molti neuroni le cui singole attività transitoriamente somma ai grandi eventi che possono essere rilevati nel potenziale di campo locale (LFP) con microelettrodi extracellulari, o nel elettroencefalogramma (EEG), il magnetoencephalogram (MEG ), e il segnale BOLD dalla risonanza magnetica funzionale (fMRI). La LFP è attualmente il metodo di scelta quando si studia l'attività neuronale popolazione con alta risoluzione temporale e spaziale alla scala mesoscopica (diverse migliaia di neuroni). Al microelettrodo extracellulare, localmente attività sincronizzata del spazialmente neighbored risultato neuroni deviazioni rapido l'LFP fino a diverse centinaia di microvolt. Quando si utilizza una serie di microelettrodi, le organizzazioni di tali deviazioni possono essere comodamente controllati nello spazio e nel tempo.
Valanghe neuronale descrivere la scala-invariante organizzazione spazio-temporale in corso di attività neuronale del cervello 2,3. Essi sono specifici per gli strati superficiali della corteccia, come stabilito in 4,5 vitro, in vivo nel ratto anestetizzato 6, e nella scimmia sveglio 7. È importante sottolineare che gli studi teorici ed empirici suggeriscono che le valanghe 2,8-10 neuronale indicano una dinamica squisitamente equilibrato stato critico della corteccia che ottimizza il trasferimento di informazioni e di elaborazione delle informazioni.
Al fine di studiare i meccanismi di sviluppo valanga neuronale, la manutenzione e la regolamentazione, in preparati in vitro sono di grande beneficio, in quanto consentono di registrazioni stabile di attività valanghiva a condizioni ben controllate. L'attuale protocollo descrive come studiare valanghe neuronali in vitro, sfruttando lo sviluppo strato superficiale in organotipica corteccia culture, culture fetta cioè, coltivate su planare, array di microelettrodi integrati (MEA, vedi anche 11-14).
Protocol
1. Sterile, Camera di vetro sigillabili con MEA per registrazioni a lungo termine
- Cilindri di vetro filettato con tappo in plastica-teflon (Asso di vetro), richiesti per la chiusura cultura sicuro e stretto da camera, vengono tagliate (Aceglass) circa 2 mm dal fondo del filetto (Fig. 1A, B). Anelli di vetro pulito da risciacquo con acqua (3x) e la bollitura per 5 minuti in 200 prove di alcol etilico, lasciare asciugare.
- Silicio soluzione aliquota tenuti ad allegare anelli di vetro alla superficie MEA. Mescolare 15 ml di parti A & B di Sylgard 184 Kit di silicone elastomero accuratamente, lasciare riposare per 15 minuti per eliminare le bolle d'aria, conservare in aliquote da 1 ml a -20 ° C.
- Colla anello di vetro alla MEA (griglia 8x8 w / elettrodo interno di terra, a 30 micron di diametro dell'elettrodo, 200/100 micron distanza inter-elettrodica di ratto / topo) (Fig. 1A, B). Raccogliere 1 ml di silicio (23 ° C) in siringa con ago piccolo calibro. Applicare del silicone di superficie di taglio grezzo di anello di vetro, vetro anello centrale su MEA, applicare un ulteriore strato di silicone intorno alla parte esterna del ring per una tenuta forte, lascia la cura per 1 - 2 ore a 60 ° C sulla piastra calda.
- Sterilizzare MEA da camera e tappi da camera in una cappa a flusso laminare da 3x risciacquo in acqua deionizzata seguita da alcool al 70% (3 x, per ultimo risciacquo lasciate riposare per 10 minuti in alcool), seguiti da 10 minuti di esposizione della camera e degli interni tappo ai raggi UV . MEA camera di autoclave (120 ° C; bagnato; min 45) e lasciare asciugare.
- Cappotto superficie MEA all'interno della camera cultura poli-D-lisina. Per i nuovi MEA, che sono piuttosto lipofile, cappotto da aspirazione goccia ripetuta della soluzione dalla griglia di elettrodi. Per MEA utilizzati, coperchio inferiore da camera con soluzione, aspirare il liquido in eccesso, far evaporare in condizioni di sterilità all'interno cappa laminare. Fissare tappo per sigillare MEA camera per la conservazione e l'uso futuro.
2. Ingredienti necessari per la preparazione e la crescita di culture organotipica
- Sciogliere agar sterile nello 0,9% di NaCl, versare in sterili Petri (Falcon, 100 x15; ~ 5 mm Livello), lasciate raffreddare e avvolgere sterile con Parafilm per la conservazione. Tagliare 20 x 10 x 5 mm da blocchi di agar solido per l'uso.
- Negozio super-colla, ad esempio Devcon Super Glue II, con la confezione pulito con 70% EtOH prima di aprire, all'interno della cappa a flusso laminare per preservare la sterilità.
- Preparare il 50% di D-glucosio (SIGMA ultra, G7528) con l'aggiunta di 40 g di glucosio per 40 ml di acqua deionizzata cultura (Sigma). Conservare in 2 ml a -20 ° C.
- Aggiungere 4 ml di 50% di D-glucosio a 500 ml di soluzione salina bilanciata Gey, e freddo per granite (miscela di liquido / cristalli di ghiaccio) in freezer prima dell'uso.
- Pollo al plasma ricostituire in 5 ml di acqua deionizzata cultura (scuotere gentile, evitare la formazione di bolle), per non accontentarsi di una soluzione 5 - 10 min, capovolgere delicatamente, e decantate il chiaro contenuto in una piastra di Petri sterile. Sterile-filtro (0,22 micron filtro pori; proteine) soluzione al plasma, aliquota 350 microlitri in cryotubes (NuncTM), conservare a -20 ° C.
- Trombina ricostituire da plasma bovino di conseguenza, sterile-filtro (0,22 micron pori del filtro), aliquota 40 microlitri in cryotubes (NuncTM), conservare a -20 ° C. Per la soluzione di lavoro, diluire 40 ml di soluzione di trombina in 375 ml di soluzione salina bilanciata Gey di w / D-glucosio.
- Preparare 400 ml di terreno di coltura mescolando 100 ml di siero di cavallo, 200 ml basale medio Aquila, 100 ml di Hank soluzione salina tamponata a cui si aggiungono 4 ml di glucosio al 50% e 2 ml di 200 mM L-glutammina. Può essere conservato 4 - 8 settimane in 100 ml bottiglie PYREX a 4 ° C.
- Preparare inibitore della mitosi, mescolando uridina 0,3 mM, 0,3 mm mm ARA-C citosina-β-D-arabinofuranoside, e 0,3 5-fluoro-2'-deossiuridina, filtro sterile, aliquota 200 ul e conservare a -20 ° C per 6 - 12 mesi.
3. Cortex e area tegmentale ventrale (VTA) dissezione tissutale (tempo: <1 ora)
- Procedura produce corteccia e sezioni di tessuto VTA per ~ 12 co-culture di ratti o topi, e viene preparato all'interno di una cappa a flusso laminare in condizioni sterili. Tempo totale per la raccolta di tessuto deve essere di 1 ora.
- Prendere sano, cuccioli ben nutriti (numero dei nati ~ 10; presenza di un 'spot latte' addominale) a 1 - 2 giorni dopo la nascita (PND). Tenere un cucciolo delicatamente il muso, lasciarlo pendere liberamente, e velocemente decapitare alla base del collo con le forbici taglienti.
- Per la rimozione del cervello, togliere la pelle (due tagli laterali forbice), taglio a forbice cranio aperto con gli occhi bene (1 sagittale taglio della linea mediana; 1 taglio coronale alla corteccia / giunzione cervelletto). Capovolgere di nuovo tutti e 4 i lembi cranio. Con una spatola affilata, tagliare frontalmente attraverso il bulbo olfattivo, spatola anticipo caudalmente sotto il cervello. Sollevare delicatamente il cervello fuori dal cranio e farla scivolare in sterili, refrigerati, Gey soluzione per un rapido raffreddamento e lo stoccaggio temporaneo. Ripetere i passaggi 3,2-3,3 per 2 cervelli più (tempo totale: <20 min).
- Per ottenere VTAtessuto, trasferire il cervello su una sterile, piatto asciutto Petri con una spatola di piccole dimensioni. Ulteriori rimuovere il liquido in eccesso facendo scorrere delicatamente ogni cervello di circa 1 cm di lato. Rimuovere tronco cerebrale da una corona, taglio verticale a livello del cervelletto con una lama di rasoio.
- Colla blocco agar per il montaggio disco per la stabilizzazione meccanica del cervello durante la procedura di taglio. Inserire una linea sottile di colla di qualche millimetro di fronte al blocco agar sul disco (evitare di toccare l'agar colla).
- Con una spatola piccola, trasferire e montare ogni cervello, polo frontale verso il basso. Assicurarsi che i poli frontali sono incollati al disco di montaggio e che le parti ventrali toccare la agar senza residui di colla in modo da ottenere la giusta stabilizzazione meccanica durante il taglio e facile decollo delle fette tagliate.
- Con attenzione immergere e sicuro disco di montaggio con montaggio cervello in un cassetto vibratome (es. Leica VT1000) pieno di sterile, refrigerati soluzione di Gey. Con una lametta accuratamente puliti (90% EtOH), tagliare fette coronali del mesencefalo a frequenza più alta vibrazione e relativamente bassa velocità di avanzamento in uno spessore di 400 500 micron. L'utilizzo di un invertito pipetta Pasteur con bulbo di aspirazione, il trasferimento e la raccolta contenente le fette VTA in 35 mm x 10 piatti pieni di Petri sterile, refrigerati soluzione Gey (Fig. 1C; vedere la targhetta anche coronale 18-20 a E22 a 15).
- Per le sezioni della corteccia, ripetere i passaggi 3,2-3,6, ma si applicano taglio verticale tra corteccia e cervelletto, e montare forebrains con polo frontale alto. Circa 3 fette coronali (350 micron di spessore) a partire a livello dello striato sono raccolti per la dissezione corteccia futuro.
- Utilizzando un micro-lama fatta di lame di rasoio rotto, sezionare ~ 2 mm di larghezza sezione coronale della corteccia frontale e delle aree del mesencefalo che contiene il VTA (Fig. 1C) sotto uno stereomicroscopio. Raccogliere separatamente sezioni di tessuto in piatti di piccole dimensioni (es.: vetrini da camera), contenente una soluzione di Gey refrigerata.
4. Montaggio Cortex e fette di tessuto VTA sul MEA (Time: <1 ora)
- Posizione MEA a temperatura ambiente al microscopio stereo con la matrice di elettrodi a fuoco. Centro a 25 gocce ml di plasma sul pulito, privo di polvere, e sterile matrice matrice di elettrodi. Utilizzando spatole piccolo, far scorrere con attenzione una corteccia e la sezione in VTA la goccia plasma.
- MEA posto sulla piastra di raffreddamento, rimettere a fuoco vista, lasciate raffreddare per circa 15 s, poi aggiungere 25 ml di trombina nel plasma goccia. Utilizzando la punta della pipetta trombina, con attenzione diffondere la plasma / trombina miscela con piccoli movimenti circolari in tutto il MEA. Non toccare l'array di elettrodi direttamente fragile. Delicatamente posizione della corteccia sull'array con bordo dorsale lungo la fila secondo elettrodo dell'array. In questo modo, gli strati superficiali sviluppo arriverà a coprire il richiamo della serie. Il VTA è messa vicino al confine ventrale della sezione corteccia (Fig. 1D).
- Cap e vagamente chiudere la camera di MEA per mantenere alto tasso di umidità, mentre il MEA / cultura montaggio si siede per ~ 5 minuti all'interno della cappa a temperatura ambiente per permettere al plasma / trombina coagulazione. Nel frattempo, ripetere il passaggio 4,1-4,3 per 3 culture più.
- Aggiungere con cautela 600 ml di terreno di coltura in piccole gocce alla camera di cultura con una siringa da 1 cc con 25 x 5 / 8 ago.
- Chiudere bene la camera e il luogo MEA MEA / montaggio cultura sul vassoio portaoggetti a dondolo all'interno dell'incubatore (Fig. 1B). Per accelerare la procedura, 3 - 4 MEA possono essere montati in sequenze che si sovrappongono. Tempo di montaggio per 12 MEA deve essere <1 ora.
- Dopo 2 giorni in vitro (DIV), aggiungere 10 ml di inibitore della mitosi. Aggiorna cultura dei media del 60% a 4 DIV e ogni 4 giorni successivi.
5. Registrazione elettrofisiologiche e generazione di stimolo
- Per stabilire la relazione tra deviazioni significative nelle potenzialità campo locale (LFP) e la tendenza dei neuroni a fuoco potenziali d'azione, dopo ~ 1 settimana 5,6 registrano l'attività spontanea a 24 kHz per circa 10 minuti da ogni elettrodo della MEA (Hardware : MEA1060 w / circuito di tranciatura, x1200 guadagno, 12 bit A / D, range 0-4096 mV, sistemi multicanale; Software: MC_Rack). Terreno è fornito attraverso l'elettrodo interno di terra, o esternamente aggiungendo un Ag / AgCl metà delle cellule.
- Separare le LFP con un filtro passa banda di 1 - 200 Hz da extracellulare attività picco (banda passante 300 - 3000 Hz). Spike attività possono essere ulteriormente classificati in attività singole e multi-unità utilizzando off-line selezionatrici picco (es. Plexon Inc.). Calcola il picco innescato medie per ogni elettrodo. Per le culture corteccia, la maggior parte medie identificherà negativo deviazioni LFP (nLFP) come momento privilegiato di spiking neuronale nella cultura.
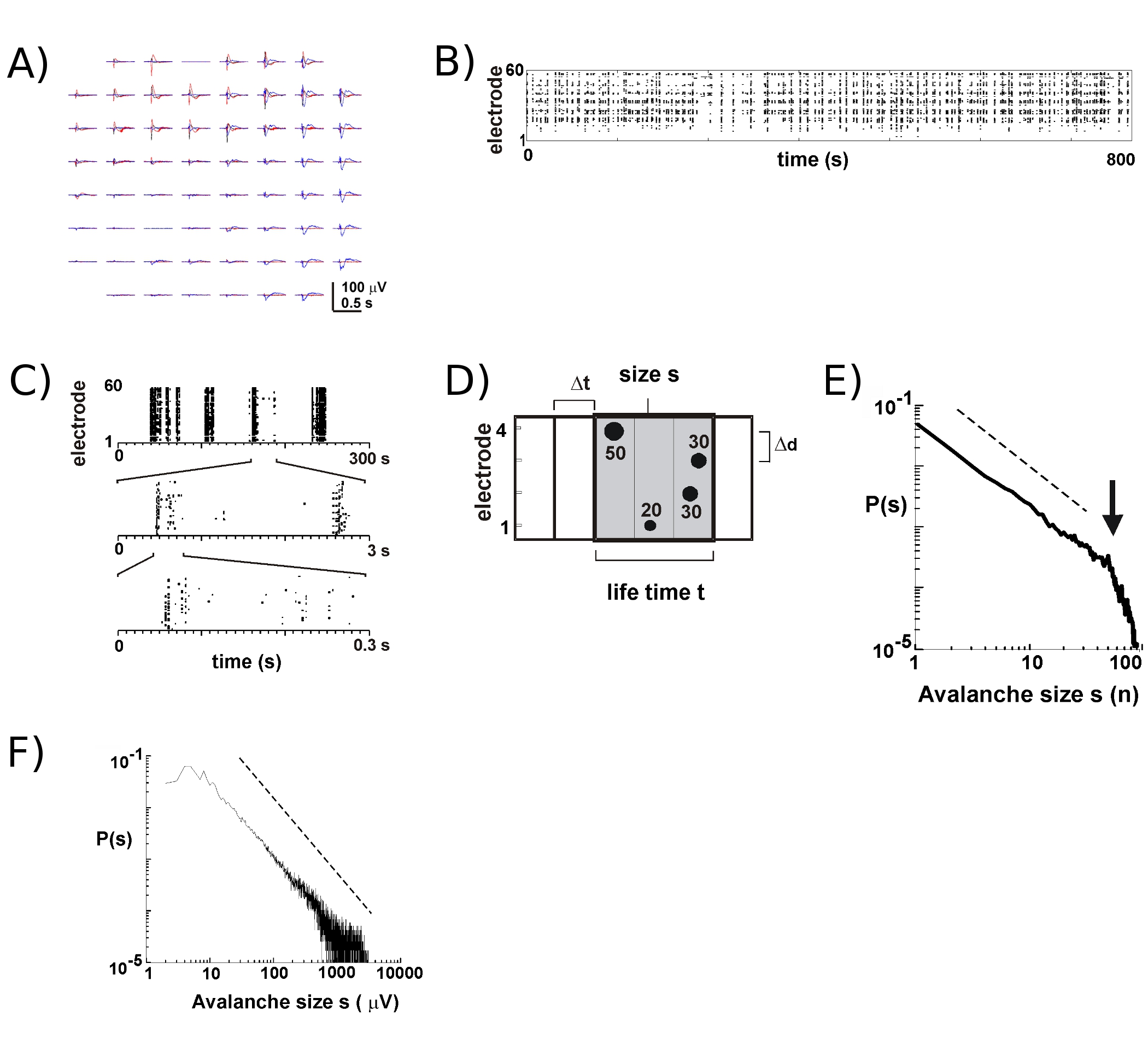
- Calcolare per ogni elettrodo una soglia di -3 deviazioni standard del rumore (SD) dalle tracce LFP (Fig. 2), Determinare i tempi e le ampiezze di picco nLFPs che attraversano soglia (Fig. 2B, C). Ha scelto un tempo di bin At (ad esempio tra 2 - 8 ms) e identificare i cluster nLFP spazio-temporale sulla matrice concatenando nLFPs da tutto elettrodi che sono gli stessi del combina tempo successivi di At lunghezza (Fig. 2D; per i dettagli vedi 2,4 , 5).
- Al fine di individuare valanghe neuronale, calcolare la dimensione di ogni cluster nLFP, ad esempio il numero di elettrodi attivi o somma delle ampiezze nLFP, costruire un istogramma dimensioni, e la trama a doppio logaritmica coordinate. Per le valanghe neuronali, la distribuzione delle dimensioni segue una legge di potenza approssimata da una linea retta in doppio logaritmica coordinate 2 (Fig. 2E, F). Vedere 16 per test statistici sulle leggi di potenza.
- Suscitare risposte evocate nel tessuto selezionando un elettrodo attraverso il quale vengono applicati corrente controllato stimoli con ampiezza S (generatore di stimoli STG 1008, i sistemi multicanale). Per ridurre i danni elettrodo, utilizzare una gamma limitata, di carica neutra stimolazione di shock singola con bipolare piazza d'onda: 50 ms con ampiezza-S seguita da 100 ms con ampiezza + S / 2 e S tra le 10-200 μA. Consultare il manuale del proprietario per ulteriori dettagli.
- Per registrare la gamma dinamica 9, stimolo registrare registrate le risposte LFP a 4 kHz frequenza di campionamento su tutti gli elettrodi dopo 500 ms dopo la stimolazione. Usa tranciatura circuito (sistemi multi-canale), che stacca la testa amplificatori palco durante la stimolazione per ridurre gli artefatti stimolo e per evitare di pre-amplificatore saturazione.
6. Rappresentante dei risultati:
Con i nuovi MEA circa 8 - 9 su 10 culture sopravvivere per molte settimane. La maggior parte dei nostri a lungo termine le registrazioni hanno luogo all'interno della incubatrice nel terreno di coltura, che ci permette di seguire lo sviluppo delle singole culture nel corso di molte settimane 5. Sulla base delle nostre esperienze, le registrazioni LFP può essere attendibilmente ottenuta con MEA utilizzato per più di 100 giorni della cultura. Al contrario, l'attività extracellulare picco è più attendibile relativamente nuovi MEA (<40 giorni cultura). In un tipico esperimento, abbiamo trasferire una MEA dal vassoio di memorizzazione (Fig. 1B, a destra) per il vassoio con la fase di testa allegata (Fig. 1B, a sinistra) tenere la camera di cultura sigillata. Per la corteccia 5, corteccia-VTA co-colture 6, così come nel ratto anestetizzato 6 e la scimmia sveglio in vivo 7, scarica neuronale durante valanghe neuronale negli strati superficiali avviene prevalentemente vicino alla deviazione di picco negativo della LFP (nLFP ). Così, l'organizzazione spazio-temporale dei gruppi neuronali sincronizzate a livello locale può essere valutata misurando la presenza di nLFPs nello spazio e nel tempo il 17 array.
Attività sul MEA tende ad emergere in cluster temporale, in modo tale che l'attività di un elettrodo è accompagnato da attività in altri siti. Forme d'onda tipica della LFP durante i periodi di tale attività sono riportati in figura 2A, da oltre tracciando 3 gruppi che si verificano alcuni secondi di distanza. Per ogni cluster, deviazioni campo negativo può essere visto più elettrodi all'interno di una finestra di 1 s. Quando si estraggono i picchi nLFP che attraversano una soglia di più negativo SD, l'attività in forma di nLFP punta è comodamente visualizzata in un raster in cui 'colonne' di punti rappresentano vicino nLFPs coincidente a vari elettrodi (Fig. 2B). L'organizzazione spazio-temporale di questa attività è piuttosto complesso; 'colonne', che appaiono più o meno omogeneo a bassa risoluzione temporale, sono composte da cluster separati a più alta risoluzione temporale e così via (fig. 2C). Infatti, l'emergere di cluster spazio-temporale nLFP è altamente organizzati in reti corticali. Più specificamente, l'organizzazione è scale-invariante per le valanghe neuronale. Ciò è dimostrato calcolando la probabilità di dimensioni dei cluster in un dato At risoluzione temporale. Qui, i cluster sono costituiti da nLFPs che si verificano negli appositi contenitori contemporaneamente o successive (Fig. 2D). Quando la dimensione di tale cluster è espressa in numero totale di nLFPs per cluster, o integrato ampiezze nLFP per cluster, le distribuzioni dei cluster dimensioni rivela una legge di potenza, la cui pendenza ha dimostrato di essere -1,5 2,4,5,7 ( fig. 2E, F). Si noti che questa distribuzione identifica una scala-invariante ordinamento delle dimensioni dei cluster che è il rapporto tra dimensioni s per KXS, dove k è un fattore costante, è -1,5 k, che è indipendente da s. Questa organizzazione legge di potenza è indipendente dalle dimensioni della matrice 2, la risoluzione temporale At 2, e la soglia utilizzata per identificare nLFP significative deviazioni 7. Perché le scale ampiezza nLFP con dimensione del gruppo neuronale 7, la scala-invariante organizzazione di nLFPs riflette una scala-invariante, cioè frattale, ordinando of gruppi neuronali sincronizzate a livello locale che comprendono tutte le dimensioni.

Figura 1. (A), viste laterali e parte superiore del MEA con ghiera filettata vetro montato, e il tappo corrispondente. (B) Vista interna del termostato. A sinistra: headstage montaggio che consenta la registrazione di una singola cultura in condizioni di incubatrice. A destra: Vassoio tenendo MEA numerosi per la crescita della cultura. Ruote laterali: motore passo-passo dispositivo controllato a dondolo per alternando fase sommerso e l'atmosfera esposti necessari per la crescita della cultura. (C) Schema per le fette di ratto coronale utilizzato per Cortex-VTA co-culture. Sezioni della corteccia (a sinistra) e le sezioni mesencefalo (al centro, a destra) contenente il VTA tegmentale ventrale (VTA, grigio) si ottengono tagliando lungo le linee tratteggiate. Ctx: corteccia; wm: sostanza bianca; cpu: striato; vta: pons: area pontina. Vedi anche piatti corrispondenti coronale 8, 18, e 20 da 15. (D) collocamento e la crescita di una singola corteccia-VTA co-culture in MEA e il suo sviluppo nei primi 9 DIV nella cultura. Nota l'appiattimento della cultura e della sua progressiva espansione sulla matrice. Parti di tessuto riflettente indicare cellule degenerate e detriti dei tessuti. Tessuto sano è opaco e grigio con transilluminazione con la luce visibile.

Figura 2. Valanghe neuronale corticale a culture organotipica. (A) Overplot di tre periodo di attività spontanea sulla matrice, separati da alcuni secondi. Si noti che ogni periodo di attività è costituito da deviazioni LFP negativo sulla elettrodi molti sul campo (ogni colore etichette un periodo di attività). (B) ore di punta negativa di nLFPs da ogni elettrodo sono assemblati in un raster di attività. 'Column'-come le strutture indicati i periodi di attività vicino a sincrono. (C) Si noti che le colonne che appaiono altamente sincronizzati in una scala di tempo costituito da più colonne alle scale temporali superiori (3 scale temporali in figura). (D) Rappresentazione schematica dell'algoritmo valanga neuronale. Su un 2 x 2 ore di punta serie di elettrodi e l'ampiezza delle deviazioni negative LFP (nLFP) attraversando una soglia di-x SD del rumore sono identificati. Organizzazione spazio-temporale di nLFPs è raggruppate in contenitori tempo successivamente attivo di At larghezza. La dimensione di un cluster è identificato sia dal numero di siti attivi, cioè con elettrodi nLFP (s = 4), o la somma integrata di ampiezze nLFP (s = 130 uV). Il tempo di vita si misura in multipli di At. (E, F) legge di potenza nella distribuzione dimensione cluster identifica i cluster come valanghe neuronale. Si noti che la scelta di una distanza interelettrodica particolare Δd per l'array (qui 200 micron) introduce una At particolare in cui la dinamica dovrebbe essere osservato. Più in particolare, il rapporto in cui Δd / At approssima la velocità media di propagazione in rete, in cui la α pendenza della legge di potenza è approssima -1,5 per valanghe neuronale 2,4,5. Si prega di cliccare qui per vedere una versione più grande della figura 2.
Discussion
1. Questioni tecniche:
- Tecnica sterile. La preparazione del MEA e preparazione culturale sono tutti eseguiti all'interno di una cappa a flusso laminare in condizioni sterili. Gli antibiotici, che influenzano l'attività neuronale, non vengono utilizzati in qualsiasi momento durante il processo di preparazione e cultura.
- Plasma / trombina coagulazione e l'aderenza del tessuto sul MEA. Tissue sopravvivenza sulla MEA richiede un attento equilibrio tra il tempo richiesto per il plasma / trombina coagulazione e il tempo di esposizione dei tessuti per l'atmosfera. Un tempo di coagulazione breve rischi prematuro distacco di sezioni dalla MEA, mentre l'esposizione prolungata in atmosfera provoca una degenerazione dei tessuti. Perché la forza della soluzione di trombina determina la velocità del processo di coagulazione, è un parametro molto importante per attaccare con successo, le culture sane in superficie MEA. Si ottengono risultati migliori con 1000 unità (1KU; 1 unità NIH = 0,324 ± 0,073 mg). Importante, incompleta miscelazione del plasma / soluzione di trombina risultati nella coagulazione spazialmente eterogeneo promozione plasma rotture durante la coltura. 'Buchi' queste plasma gravemente nuocere alla salute della cultura e staccare la cultura dal microelettrodi, compromettendo così la qualità della registrazione elettrofisiologica. Lavorare con piastre di raffreddamento durante la MEA / tessuto assemblaggio rallenta la coagulazione e consente la miscelazione omogenea del plasma / soluzione di trombina e il posizionamento corretto delle sezioni di tessuto.
Allo stesso modo, con l'aggiunta di medie gocce solo alla camera di cultura di sommergere la cultura dopo soli 5 minuti di coagulazione, riduce notevolmente il rischio di distacchi del tessuto a causa di tensioni superficiali. Una cultura di successo si appiattisce sulla MEA e leggermente ampliare mostrando una crescita sana per settimane, senza alcun segno importante di incompleta tessuto-MEA contatto o degenerazione dei tessuti (es. fig. 1D). - Dissezione dei tessuti. Coltelli Micro hanno notevolmente migliorato il nostro processo di dissezione. Noi usiamo due lame di rasoio bordo (Strumenti Belle Scienza - fragili lame di bisturi - 100050-00), da cui abbiamo diviso ~ 2 mm di larghezza 'lame' con una pinza, e tenerli con un supporto bisturi. Campioni di tessuto sezionato dalla fetta coronale con una superficie liscia, movimento verticale fermo della lama quindi riducendo notevolmente lo stress meccanico a causa della trazione del tessuto complessivo miglioramento della salute della cultura.
- Tessuto di raffreddamento. Il corretto raffreddamento delle fette e sezioni di tessuto durante la preparazione è essenziale per il successo della cultura. Noi usiamo piatti freddi su misura costruito su elementi Peltier attaccato sotto ad un disco di metallo. Il calore prodotto dall'elemento Peltier viene rimosso attraverso la perfusione acqua fredda. Questo riduce notevolmente i tempi di preparazione e standardizza il raffreddamento durante ogni fase della preparazione (Labs Dold & Engineering 131 Plantation Dr. Seguin, TX 78155;. (830) 560-1471)).
- Condizione incubatrice. Una misura incubatore con precise condizioni a dondolo interno è stato fondamentale per il nostro successo nel coltivare le fette su MEA. Sulla base di un originale in-house di progettazione Multichannelsystems (attualmente non disponibile in commercio), il dispositivo a dondolo interno è costituito da vassoi che sono collegati a due ruote laterali di grandi dimensioni. Motori passo-passo e controlli computerizzati consentono una precisa traiettoria a dondolo (angolo a dondolo, dondolo velocità e intermittenti pause). In ultima analisi, culture fetta devono essere esposti a medio ambiente e la cultura in alternanza lento. L'approccio tradizionale è quello di posizionare le culture fetta in tubi stretti che ruotano lentamente lungo ci asse più lungo. Qui, lenta rotazione non produce sollecitazioni meccaniche dovute alla stessa rotazione e la velocità di rotazione è abbastanza alta da ottenere un ottimale 'alimentazione / respirare' ciclo di circa 5 - durata 10 min. L'interno, più compatto della camera di MEA, il suo volume totale di circa 2 cc, e il piccolo volume di terreno di coltura richiesta per il condizionamento di media dal tessuto stesso, pone una sfida significativa. Facendo scorrere l'MEA tra ± ~ 70 ° angolo (tempo di ciclo: ~ 200 s), rallentando la velocità a dondolo come le transizioni cultura tra la fase liquida e l'atmosfera, e ad arrestare la dondolo agli angoli estremi per la prolungata esposizione all'atmosfera è stato essenziale per la sopravvivenza della cultura.
2. Età evolutiva di culture corteccia per studiare valanghe neuronale
Fette acuta da corteccia di ratto sono comunemente prese a PND 0-1 e colto per molte settimane sulla MEA. I primi studi hanno chiaramente dimostrato che singole culture fetta corteccia, dopo diverse settimane in vitro, mantenere una struttura a strati con tipi di cellule identificabili che possono essere facilmente rispetto alle classi di cellule in vivo 18,18-21. L'organizzazione a strati in questo sistema in vitro è stato convenientemente utilizzata per lo studio del talamoinnervazione della corteccia durante lo sviluppo 22-24, così come per la guida delle strutture sottocorticali, come lo striato 25,26. In effetti, la specificità nella formazione di connessioni neuronali all'interno e tra regioni del cervello permette per la costruzione di complessi sistemi in vitro che riprendono numerosi sistemi di proiezione dettagliate, ad esempio quello della corteccia, dei gangli basali circuiti 27-30.
Dopo 4 - 6 settimane in vitro, solo fette corteccia 31 e fettine di corteccia co-coltura con striato 26 o 32 talamo manifestazione spontanea e GIÙ stati tipicamente presenti in vivo sul ratto anestetizzato uretano 33. L'organizzazione bel temporale di questi Stati-up porta il marchio di garanzia di annidati θ e γ-oscillazioni indicativo di una rete elettrofisiologico maturo di neuroni piramidali e interneuroni GABAergici fast-spiking 31. È importante sottolineare che, in assenza di recettori della dopamina D2-stimolazione, la maturazione degli interneuroni corticali parvalbumina-positivi è ritardata di circa 2 settimane nelle culture fetta corteccia 34. In linea con questi risultati, la durata dello sviluppo di annidati θ-, β-e γ-oscillazioni corrisponde a quella in vivo, quando fette corteccia sono co-coltura con l'area tegmentale ventrale (VTA), che contiene i neuroni dopaminergici che proiettano la corteccia 6.
Questi studi indicano che quando si studia valanghe neuronale, che dipendono essenzialmente dal maturo inibizione veloce GABAergici e si trovano negli strati superficiali della corteccia 2,4, grande cura deve essere presa per assicurare corretta maturazione del tessuto corticale. Mentre valanghe neuronale sorgono nelle culture corteccia singolo nel corso di tempo di 2 - 5 settimane 4, quando si richiede un corso evolutivo tempo che è abbinato al vivo nello sviluppo, fette corteccia bisogno di un appropriato stimolazione del recettore della dopamina, ad esempio di co-coltura fette corteccia con la VTA 6.
Disclosures
Nessun conflitto di interessi dichiarati.
Acknowledgments
Questo studio è stato finanziato dalla Divisione del programma di ricerca intramurale (DIRP) del National Institute of Mental Health, National Institutes of Health.
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}