

Summary
En robust sätt att studera neuronala laviner, dvs skala-invarianta tid och rum aktivitet skurar, ett tecken på kritiskt läge dynamik i cortex. Laviner uppstår spontant i att utveckla ytliga lager av odlade hjärnbarken som möjliggör långsiktiga mätningar av verksamheten med plana integrerad multi-elektrod kedjor (MEA) under exakt kontrollerade förhållanden.
Abstract
Cortex är spontant aktiva, även i avsaknad av en viss insatsvara eller motoreffekt. Under utveckling är denna verksamhet viktig för migration och differentiering av hjärnbarken celltyper och bildandet av neuronala kontakter 1. I den mogna djuret, speglar pågående aktivitet det förflutna och det nuvarande tillståndet av ett djur där sensoriska stimuli är sömlöst integrerade för att beräkna framtida åtgärder. Således är en klar förståelse för organisationen av pågående dvs spontan aktivitet en förutsättning för att förstå cortex funktion.
Numerous inspelningsteknik visade att pågående aktivitet i cortex består av många nervceller whose enskilda aktiviteter transiently summa till större evenemang som kan upptäckas i den lokala området potential (LFP) med extracellulärt microelectrodes eller i elektroencefalogram (EEG), den magnetoencephalogram (MEG ), och de djärva signalen från funktionell magnetresonanstomografi (fMRI). Den LFP är för närvarande den metod som föredras när man studerar neuronala befolkningen aktivitet med hög temporal och spatial upplösning på mesoskopisk skala (flera tusen nervceller). På den extracellulära mikroelektrod, lokalt synkroniserad verksamhet rumsligt neighbored nervceller resultera i snabb avvikelser i LFP upp till flera hundra mikrovolt. När du använder en rad microelectrodes, kan organisationer sådana omläggningar vara bekvämt övervakas i tid och rum.
Neuronala laviner beskriva skala invarianta Spatiotemporal organisation av pågående neuronal aktivitet i hjärnan 2,3. De är specifika för de ytliga lagren av hjärnbarken som fastställts in vitro 4,5, in vivo i sövda råttor 6, och i vaken apa 7. Viktigt både teoretiska och empiriska studier 2,8-10 tyder på att neuronala laviner visar en utsökt balanserad kritiskt tillstånd dynamiken i hjärnbarken som optimerar informationsöverföring och informationsbehandling.
För att studera mekanismerna av neuronala lavin utveckling, underhåll och reglering, in vitro-preparat är till stor nytta, eftersom de möjliggör stabila inspelningar av lavin verksamhet under exakt kontrollerade förhållanden. Det nuvarande protokollet beskriver hur man studera neuronala laviner in vitro genom att utnyttja ytliga lagret utveckling i organotypic cortex kulturer, dvs kulturer skiva, som odlas på plan, integrerade mikroelektrod arrayer (MEA, se även 11-14).
Protocol
1. Sterila, förseglad Glas avdelningen med MEA för långsiktigt Inspelningar
- Gängade glascylindrar med Teflon-plastlock (Ace Glas), som krävs för säker och tät kultur kammare stängning, skärs (Aceglass) ca 2 mm från botten av tråden (Fig. 1A, B). Rengör glas-ringar genom att skölja med vatten (3x) och koka i 5 min i 200 bevis etyl-alkohol, låt torka.
- Alikvotera kisel lösning som krävs för att fästa glas ringar för att MEA yta. Blanda 15 ml av delarna A & B i Sylgard 184 Silicone Elastomer Kit ordentligt, låt sitta i 15 min att ta bort luftbubblor, lagra i 1 ml alikvoter vid -20 ° C.
- Limma glas ringen MEA (8x8 rutnät w / interna jordelektroden, 30 ìm elektrod diameter, 200/100 ìm mellan elektrod avstånd för råtta / mus) (Fig. 1A, B). Ta upp 1 ml av kisel (23 ° C) i spruta med små gauge nål. Applicera silikon till opolerade snittyta av glas ring, mitt glas ringen på MEA, applicera ett extra lager av kisel runt utsidan av ringen för en starkare tätning, låt botemedel för 1 - 2 timmar vid ~ 60 ° C på värmeplatta.
- Sterilisera MEA kammare och kammarmusiker mössor i ett laminärt flöde huva med 3x skölj i avjoniserat vatten följt av 70% alkohol (3 x, för sista sköljningen låt sitta i 10 min i alkohol), följt av 10 min exponering av kammare och mössa inredning för UV-ljus . Autoklav MEA kammare (120 ° C, våt, 45 min) och låt torka.
- Coat MEA ytan inne kultur kammare med poly-D-lysin. För nya multilaterala miljöavtal, som är ganska lipofila, päls av upprepade droppe aspiration av lösningen från elektroden nätet. För används MEA, täck kammare botten med lösningen, aspirera överflödig vätska och låt det avdunsta under sterila förhållanden inne laminära huva. Sätt lock att täta MEA kammare för lagring och framtida användning.
2. Ingredienser krävs för att utarbeta och tillväxt Organotypic kulturer
- Lös steril agar i 0,9% NaCl, häll steril petriskål (Falcon, 100 x15, ~ 5 mm nivå), låt svalna och sterila wrap med Parafilm för förvaring. Cut 20 x 10 x 5 mm kvarter från fast agar för användning.
- Store superlim, t.ex. Devcon superlim II, med förpackningen torkas av med 70% EtOH innan de öppnas, innanför laminär strömning huven för att bevara sterilitet.
- Förbered 50% D-Glukos (SIGMA ultra, G7528) genom tillsats av 40 g glukos till 40 ml avjoniserat kultur vatten (Sigma). Förvaras i 2 ml alikvoter vid -20 ° C.
- Tillsätt 4 ml av 50% D-glukos till 500 ml Gey Balanced Salt lösning och kyla till slask (blandning av flytande / iskristaller) i frysen före användning.
- Bered kyckling plasma i 5 ml avjoniserat kultur vatten (skaka försiktigt, undvika bildning av bubblor), låt lösning nöja för 5 - 10 min, snurra försiktigt och decantate det klara innehållet i en steril petriskål. Sterila-filter (0,22 ìm por-filter, protein) plasma-lösning, alikvot 350 l till cryotubes (NuncTM), förvara vid -20 ° C.
- Bered trombin från nötkreatur plasma därmed steril-filter (0,22 ìm pore filter), alikvot 40 l till cryotubes (NuncTM), förvara vid -20 ° C. För fungerande lösning, späd 40 ìl av trombin lösning i 375 ìl Gey balanserade saltlösning W / D-glukos.
- Förbered 400 ml odlingsmedium genom att blanda 100 ml av häst serum, 200 ml bassubstratets Eagle, 100 ml Hanks saltlösning som 4 ml 50% glukos och 2 ml av 200 mM L-glutamin tillkommer. Kan lagras 4 - 8 veckor i 100 ml PYREX flaskor vid 4 ° C.
- Förbered mitos hämmare genom att blanda 0,3 mM uridin, 0,3 mM Ara-C cytosin-β-D-arabinofuranoside, och 0,3 mm 5-fluoro-2'-deoxiuridin, sterila filter, alikvot 200 l och förvara vid -20 ° C i 6 - 12 månader.
3. Cortex och ventrala tegmentumområdet (VTA) Tissue Dissection (tid: <1 timme)
- Förfarande ger cortex och VTA sektioner vävnad för ~ 12 co-kulturer från råttor eller möss, och är berett i ett laminärt flöde huva under sterila förhållanden. Total tid för mjukpapper uppsamling bör än 1 tim.
- Ta friska, välnärda ungar (kullstorlek ~ 10, närvaron av en buken "mjölk spot") vid 1 till 2 postnatal dagar (PND). Håll en valp försiktigt genom nosen, låt det hänga fritt och snabbt halshugga vid basen av halsen med vassa sax.
- För hjärnan bort, ta bort hud (två laterala sax klipper), skär skallen öppna med fina ögon sax (1 sagital mittlinjen skära, 1 koronalt skuret i hjärnbarken / lillhjärnan korsningen). Vänd tillbaka alla 4 skallen klaffar. Med en skärpt spatel, skär frontalt genom luktbulben, avancera spatel caudally under hjärnan. Lyft försiktigt hjärnan ur skallen och låter den glida in i sterila, kylda, Gey lösning för snabb kylning och tillfällig förvaring. Upprepa steg från 3,2 till 3,3 för 2 fler hjärnor (total tid: <20 min).
- För att få VTAvävnad, överförs hjärnan på en steril, torr petriskål med en liten spatel. Ytterligare bort överflödig vätska genom att försiktigt dra varje hjärna ca 1 cm i sidled. Ta bort hjärnstammen genom en koronalt, vertikal klippa i nivå med lillhjärnan med ett rakblad.
- Limma agar blocket på montering skivan för mekanisk stabilisering av hjärnor under skivning förfarande. Lägg en tunn linje av superlim några millimeter framför den agar blocket på disken (undvik lim röra agar).
- Använd en liten spatel, överföring och montera varje hjärna, främre stolpen ner. Se till att främre stolpar limmas till montering skiva och att ventrala sidorna vidrör agar utan limrester i syfte att uppnå god mekanisk stabilisering under sågning och lätt lättning av skurna skivor.
- Försiktigt dyka och säker montering skiva med hjärnan montering i en vibratome fack (t.ex. Leica VT1000) fylld med steril, kylda Gey lösning. Med en noga rengjord rakblad (90% EtOH), skär coronal skivor av mellanhjärnan på högsta vibrationsfrekvens och relativt låga fart framåt med en tjocklek på 400 500 ìm. Använda en inverterad pasteurpipett med sugboll, överföra och samla skivor som innehåller VTA i 35 x 10 mm petriskålar fyllda med sterilt, kylda Gey lösning (figur 1C, se även koronalt platta från 18 till 20 på E22 i 15).
- För cortex sektioner, upprepa steg från 3,2 till 3,6, men gäller vertikala snitt mellan hjärnbarken och lillhjärnan, och montera forebrains med främre stolpe upp. Cirka 3 koronalt skivor (350 ìm tjocklek) börjar på samma nivå som striatum samlas för framtida cortex dissekering.
- Med hjälp av en mikro-kniv är gjord av krossade rakblad, dissekera ~ 2 mm bred koronalt avsnitt av frontala cortex och områden mellanhjärnan som innehåller VTA (bild 1C) under ett stereomikroskop. Samla vävnadssnitt separat i små rätter (t.ex. kammare diabilder) fylld med kylda Gey lösning.
4. Montering Cortex och VTA Slices Tissue på MEA (Tid: <1 tim)
- Position MEA i rumstemperatur under en lupp med elektrodbäraren i fokus. Center en 25 l droppe plasma på den rena, dammfri och steril elektrodbäraren matris. Med små spatulae, skjut försiktigt ett cortex och VTA avsnitt i plasma droppen.
- Placera MEA om kylplatta, omfokusera visa, låt kyla för ~ 15 sekunder, lägg sedan till 25 ìl av trombin i plasma droppen. Med hjälp av trombin pipettspetsen försiktigt sprida plasma / trombin blandning med små cirkelrörelser över hela MEA. Rör inte spröda elektrodbäraren direkt. Försiktigt placera cortex på matrisen med rygg-kant längs den andra elektroden raden i matrisen. På så sätt kommer utvecklingsländerna ytliga lager täcker så småningom påminnelse om arrayen. Den VTA är placerad intill den ventrala gränsen av hjärnbarken avsnitt (bild 1D).
- Cap och löst stänga MEA kammaren att behålla hög luftfuktighet medan MEA / kultur montering sitter för ~ 5 min inne i huvan i rumstemperatur för att möjliggöra för plasma / trombin koagulering. Samtidigt upprepar steg från 4,1 till 4,3 till 3 fler kulturer.
- Tillsätt försiktigt 600 l odlingsmedium i små droppar med den kultur kammaren med en 1 cc spruta med 25 x 5 / 8 nål.
- Tätt nära MEA kammaren och placera MEA / kultur montering på gungande avställningsplats inne i inkubatorn (Fig. 1B). För att påskynda förfarandet, 3 till 4 MEA kan monteras i överlappande sekvenser. Monteringstiden för 12 miljöavtal bör vara <1 timme.
- Efter 2 dagar in vitro (DIV), tillsätt 10 ìl av mitosen hämmare. Uppdatera närsubstrat med 60% vid 4 DIV och var 4 dagar därefter.
5. Elektrofysiologiska Inspelning och stimulans Generation
- Att fastställa förhållandet mellan betydande avvikelser i det lokala området potential (LFP) och tendensen av nervceller till brand aktionspotentialer, efter ~ 1 vecka 5,6 rekord spontan aktivitet vid 24 kHz för ~ 10 min från varje elektrod av MEA (Hardware : MEA1060 w / blanking krets, X1200 förstärkning, 12 bitars A / D, intervall 0-4096 mV, flerkanaliga system, Programvara: MC_Rack). Ground tillhandahålls antingen via den interna jordelektroden, eller externt genom att lägga till en Ag / AgCl-halv cell.
- Separera LFP med ett bandpassfilter av 1 - 200 Hz från extracellulära spik aktivitet (bandpass 300 - 3000 Hz). Spike aktivitet kan ytterligare delas in i enkel-och multi-enhetens verksamhet med hjälp av off-line spik sorterare (t.ex. Plexon Inc.). Beräkna spik utlöste medelvärden för varje elektrod. För cortex kulturer, kommer de flesta medelvärden identifiera negativa LFP nedböjningar (nLFP) som det lämpligaste tiden för neuronala tillsatta i kulturen.
- Beräkna för varje elektrod en tröskel på -3 standardavvikelser för buller (SD) från LFP spår (Fig. 2)Fastställa rusningstrafik och amplitud av nLFPs som korsar gränsen (Fig. 2B, C). Välj ett Dt tid bin (t ex mellan 2 - 8 ms) och identifiera Spatiotemporal nLFP kluster på matrisen genom att sammanfoga nLFPs från alla elektrod som är i samma successiva gång kombinerar längd Dt (Fig. 2D, för detaljer se 2,4 , 5).
- För att identifiera neuronala laviner, beräkna storleken på varje nLFP kluster, exempelvis antalet aktiva elektroder eller summan av nLFP amplituder, konstruera en storlek histogram och tomt i dubbel logaritmisk koordinater. För neuronala laviner följer storleksfördelning en makt lag approximeras med en rak linje i dubbel logaritmisk koordinater 2 (Fig. 2E, F). Se 16 för statistiska test på strömmen lagar.
- Framkalla evoked potential i vävnaden genom att välja en elektrod där strömstyrda stimuli med amplituden S tillämpas (Stimulus generator STG 1008, Multichannel Systems). För att minska elektrod skada, använda en rad begränsad, charge-neutral stimulering av enstaka stötar med bipolär kvadrat vågform: 50 ìs med amplitud-S följt av 100 ìs med amplitud + S / 2 och S mellan 10 till 200 μA. Se bruksanvisningen för ytterligare information.
- För att spela in det dynamiska omfånget 9, spela stimulans inspelade LFP svar vid 4 kHz samplingsfrekvens på alla elektroder efter 500 ms efter stimulering. Använd blanking kretsar (Multi-channel system) som bryter den förstärkare huvudet scenen under stimulering för att minska stimulans artefakter och förhindra förförstärkare mättnad.
6. Representativa resultat:
Med nya MEA ca 8 - 9 av 10 kulturer kommer att överleva i flera veckor. De flesta av våra långsiktiga inspelningarna äger rum inne i inkubatorn i ett odlingsmedium som tillåter oss att följa utvecklingen av enskilda kulturer under loppet av många veckor 5. Baserat på våra experiment, kan LFP inspelningar på ett tillförlitligt sätt erhållas med multilaterala miljöavtal används för mer än 100 kultur dagar. Däremot är extracellulära topp aktivitet mer tillförlitligt sätt med relativt nya multilaterala miljöavtal (<40 kulturen dagar). I ett typiskt experiment, överför vi en MEA från lagring brickan (Fig. 1B, till höger) att facket med huvudet stadium monterat (bild 1B, vänster) att hålla kulturen kammaren förseglas. För cortex 5, Cortex-VTA co-kulturer 6, liksom i den sövda råttor 6 och vakna apan in vivo 7, neuronaktivitet under nervsystemets laviner i ytliga lager sker främst nära toppen negativa böjningen av LFP (nLFP ). Därmed kan Spatiotemporal organisation av lokalt synkroniseras neuronala grupper beräknas genom att mäta förekomsten av nLFPs i tid och rum på matrisen 17.
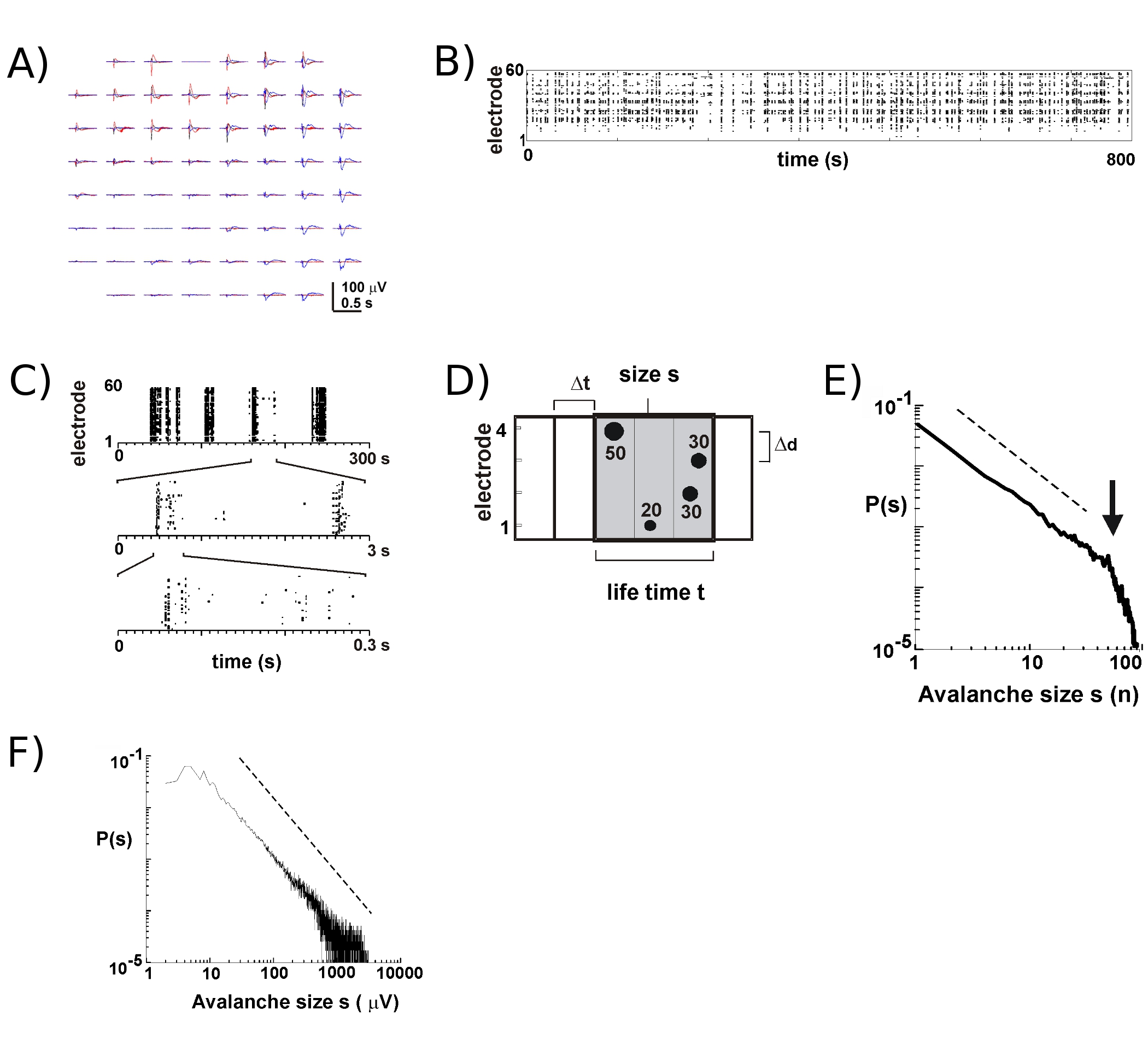
Aktivitet på MEA tenderar att dyka upp i timliga kluster, är sådan att verksamheten på en elektrod tillsammans med aktivitet på andra sajter. Typiska vågformer av LFP under sådan verksamhet perioder visas i figur 2A, med över plottning 3 kluster uppstår några sekunders mellanrum. För varje kluster kan negativa fält omläggning ses på flera elektroder i ett fönster på 1 s. Vid utvinning nLFP toppar som passerar en tröskel på flera negativa SD, är aktiviteten i form av nLFP rusningstid enkelt visualiseras i ett raster där "kolumner" av prickar representerar nära sammanfallande nLFPs på olika elektroder (Fig. 2B). Den Spatiotemporal organisation av denna verksamhet är ganska komplext, "kolumner", som visas mer eller mindre homogen med låg tidsupplösning, består av separata grupper med högre tidsupplösning och så vidare (Fig. 2C). I själva verket är uppkomsten av Spatiotemporal nLFP kluster välorganiserad i kortikala nätverk. Mer specifikt är organisationen skala invariant för neuronala laviner. Detta visas av att beräkna sannolikheten för klusterstorlekar vid en viss temporal upplösning Dt. Här är kluster består av nLFPs som förekommer i samma eller efterföljande gång kärl (Fig. 2D). När storleken på ett sådant kluster är uttryckt i totalt antal nLFPs per kluster, eller integrerade nLFP amplituder per kluster, avslöjar klustret storlek distributioner en makt lag, vars lutning har visat sig vara -1,5 2,4,5,7 ( Fig.. 2E, F). Observera att denna distribution identifierar en skala invariant beställning av kluster storlekar som är förhållandet mellan storlek s till kxs, där k är en konstant faktor, k -1,5, som är oberoende av S. Denna makt lag organisation är oberoende av storleken på arrayen 2, nedböjningar den temporala upplösningen Dt 2, och tröskeln för att identifiera betydande nLFP 7. Eftersom nLFP amplitud skalor med neuronala gruppstorlek 7, återspeglar omfattningen-invarianta organisation nLFPs en skala invariant, dvs fraktal, beställning of lokalt synkroniseras neuronala grupper som inkluderar alla storlekar.

Figur 1. (A) Sida och topp utsikt över MEA med gängad glas ringen monteras, och motsvarande mössa. (B) Insidan av inkubatorn. Vänster: headstage fäste möjliggör inspelning från en enda kultur i kuvös skick. Höger: fack håller många MEA för kultur tillväxt. Side hjul: stegmotor styrd gungande enhet för växelström dränkt och atmosfär-exponerade fas krävs för kultur tillväxt. (C) Schematisk ritning för koronala råtta skivor används för Cortex-VTA co-kulturer. Cortex sektioner (vänster) och delar mitthjärnan (mitten, höger) innehåller VTA ventrala tegmentumområdet (VTA, grå) erhålls genom att skära längs de streckade linjerna. Ctx: cortex, WM: vit substans; cpu: striatum; VTA: Pons: Pontine området. Se även motsvarande koronalt tallrikar 8, 18 och 20 med 15. (D) Placering och tillväxt av en enda Cortex-VTA co-kulturer på MEA och dess utveckling under de första 9 DIV i kultur. Notera flackare kulturen och dess gradvis utbyggnad på matrisen. Reflekterande vävnad delar visar degenererade celler och vävnader skräp. Frisk vävnad är ogenomskinlig och gråaktig i genomlysning med synligt ljus.

Figur 2. Neuronal laviner i kortikala organotypic kulturer. (A) Overplot tre period av spontan aktivitet på matrisen, separerade med flera sekunder. Observera att varje verksamhet period består av negativa LFP nedböjningar på många elektroder på matrisen (varje färg etiketter en aktivitet perioden). (B) Negativa högtrafik av nLFPs från varje elektrod monteras till ett raster av aktivitet. "Column'-liknande strukturer angivna perioder av nära synkron aktivitet. (C) Observera att kolumner som mycket visas synkroniserad med en tidsskala består av flera kolumner på högre tidsmässiga skalor (3 tidsmässiga skalor visas). (D) Schematisk representation av neuronala lavin algoritm. På en 2 x 2 elektrodbäraren topp tid och amplitud av negativa LFP nedböjningar (nLFP) korsar en gräns på-x SD för buller identifieras. Spatiotemporal organisation nLFPs är klustrade i successivt aktiv tid lådor i bredd Dt. Storleken på ett kluster identifieras av antingen antalet aktiva sajter, dvs elektroder med nLFP (s = 4), eller den integrerade summan av nLFP amplituder (s = 130 μV). Den livslängd mäts i multiplar av Dt. (E, F) Kraft lag i kluster storleksfördelning identifierar kluster neuronala laviner. Notera att valet av en viss interelectrode avstånd Δd för matrisen (här 200 mikrometer) införs en viss Dt där dynamiken bör observeras. Mer specifikt förhållandet där Δd / Dt ungefär den genomsnittliga utbredning hastigheten i nätverket, där lutningen α av kraften lagen är ungefär -1,5 för neuronala laviner 2,4,5. Vänligen klicka här för att se en större version av figur 2.
Discussion
1. Tekniska frågor:
- Steril teknik. Beredningen av multilaterala miljöavtal och kultur förberedelser är alla utförda i ett laminärt flöde huva under sterila förhållanden. Antibiotika, som påverkar nervaktivitet, används inte vid något tillfälle under förberedelserna och kultur process.
- Plasma / trombin koagulation och vävnad anslutning på MEA. Tissue överlevnad på MEA kräver en noggrann avvägning mellan den tid som krävs för plasma / trombin koagulation och tiden av vävnad utsätts för atmosfären. En kort koagulationstid risk för tidig avlossning av sektioner från MEA, medan långvarig exponering för atmosfär utlöser vävnad degeneration. Eftersom styrkan i trombin lösningen bestämmer hastigheten på koagulationen processen är det en mycket viktig parameter för ett framgångsrikt fästa, friska kulturer att MEA ytan. Vi får bästa resultat med 1000 enheter (1KU, 1 NIH enhet = 0,324 ± 0,073 mg). Viktigt är ofullständig blandning av plasma / trombin lösning resulterar i rumsligt heterogena koagulation främja plasma-spricker under odling. Dessa plasma "hål" allvarligt påverka kultur hälsa och lossa kulturen från microelectrodes och därmed äventyra kvaliteten på elektrofysiologiska inspelningen. Arbeta med kyla plattor under MEA / vävnad montering bromsar koagulation och tillåter homogen blandning av plasma / trombin lösning och korrekt placering av vävnadssnitt.
På samma sätt, genom att lägga medel i enstaka droppar med den kultur kammaren att dränka den kultur efter bara 5 minuter av koagulation, kraftigt minskar risken för vävnaden avdelningar på grund av ytan spänningar. En framgångsrik kultur planar på MEA och något utöka visar god tillväxt i veckor, utan några större tecken på ofullständig vävnadsspecifika MEA kontakt eller vävnad degeneration (t.ex. bild. 1D). - Tissue dissekering. Micro knivar har klart förbättrat vår dissektion process. Vi använder dubbla blad kanten rakblad (Fine Science Tools - krossbart skalpell blad - från 100.050 till 00), som vi delade ~ 2 mm breda "blad" med tång, och hålla dem med en skalpell hållare. Vävnadsprover dissekerade från koronala segment med hjälp av en slät, vertikal bestämd rörelse på bladet vilket avsevärt minskar mekanisk stress på grund av att dra av vävnaden övergripande förbättra kultur hälsa.
- Tissue kylning. Ordentlig kylning av skivor och sektioner vävnad under förberedelserna är viktigt för kulturen framgång. Vi använder skräddarsydda kalla plattor byggt av Peltier-element fäst under för att en metall skiva. Den värme som produceras av Peltier elementet avlägsnas genom kallt vatten perfusion. Detta reducerar kraftigt förberedelsetiden och standardiserar kylning under varje skede av beredningen (Dold Labs & Engineering 131 Plantation Dr Seguin, TX 78.155;. (830) 560-1471)).
- Inkubator skick. En skräddarsydd inkubator med exakt interna gungande förhållanden har varit avgörande för vår framgång i att odla skivor på MEA. Baserat på ett original i egen design av Multichannelsystems (för närvarande inte kommersiellt tillgängliga), består den interna gunga produkt av brickor som är knutna till två stora sidan hjul. Stegmotorer och datoriserade kontroller möjliggör en exakt gungande bana (gunga vinkel, vaggande hastighet och intermittent pauser). Ytterst slice kulturer måste utsättas för stämning och substratet i långsam växling. Den traditionella metoden är att placera skiva kulturer i smala rör som sakta roterar längs det längsta axel. Här, producerar långsamma rotation inte mekanisk stress på grund av rotation i sig, och rotationen hastigheten är tillräckligt hög för att uppnå en optimal "utfodring / andning"-cykel på cirka 5 - 10 min varaktighet. Den mer kompakta inre av MEA kammaren, den totala volymen av ~ 2 cc, och de små kultur medelhög volym för media konditionering av vävnaden i sig, innebär en stor utmaning. Genom att gunga MEA mellan ± ~ 70 ° vinkel (cykeltid: ~ 200 s), bromsa den gungande hastigheten som den kultur övergångar mellan vätskefasen och atmosfär, och stoppa gunga vid extrema vinklar för utökad exponering i atmosfären har varit viktigt för kultur överlevnad.
2. Developmental ålder av hjärnbarken kulturer för att studera neuronala laviner
Akut skivor från råtta cortex ofta fattas på PND från 0 till 1 och odlade för många veckor på MEA. Tidiga studier har tydligt visat att ensamstående cortex skiva kulturer, efter flera veckor in vitro, bibehålla en skiktad struktur med identifierbara celltyper som lätt kan jämföras med in vivo-cell klasser 18,18-21. Den skiktade organisation i detta in vitro-system har varit bekvämt användas för att studera thalamicinnervation av hjärnbarken under utveckling 22-24, samt för att driva subkortikala strukturer såsom striatum 25,26. I själva verket tillåter specificitet i bildandet av neuronala kontakter inom och mellan områden i hjärnan för byggande av komplexa in vitro-system som återerövra många detaljerade projektion system, t.ex. den i hjärnbarken, basala ganglierna kretsar 27-30.
Efter 4 - 6 veckor in vitro enda cortex skivor 31 och cortex skivor co-odlade med striatum 26 eller thalamus 32 Visa spontana upp-och ner stater som normalt återfinns i vivo i uretan sövda råttor 33. Den fina tidsmässiga organisationen av dessa upp-stater bär ett kännetecken för kapslade θ-och γ-svängningar tyder på en elektrofysiologiskt mogen nätverk av pyramidala nervceller och snabbt standardtillsatser GABAergic interneuronen 31. Viktigt i brist på dopamin D2-receptorn stimulans, är mognaden av parvalbumin-positiva kortikala interneuron försenad med ca 2 veckor i hjärnbarken slice kulturer 34. I linje med dessa slutsatser, den utvecklande tid under kapslade θ-, β-och γ-svängningar matchas till det in vivo när cortex skivor är co-odlade med ventrala tegmentumområdet (VTA), som innehåller dopaminerga neuron projicerar till cortex 6.
Dessa studier visar att när man studerar neuronala laviner, som ytterst beror på mogna snabbt GABAergic hämning och finns i ytliga lagren av hjärnbarken 2,4, har stor omsorg skall vidtas för att säkerställa korrekt mognad av kortikal vävnad. Medan neuronala laviner uppstår i enstaka cortex kulturer under tidsförloppet av 2 - 5 veckor 4, då kräver en utvecklande tid kurs som är anpassad till in vivo-utveckling, cortex skivor behöver ett lämpligt dopamin-receptorn stimulans, t.ex. genom samtidig odling cortex skivor med VTA 6.
Disclosures
Inga intressekonflikter deklareras.
Acknowledgments
Denna studie har finansierats av avdelningen för Interna forskningsprogram (DIRP) av National Institute of Mental Health, National Institutes of Health.
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}