

Summary
Korteks kritik devlet dinamikleri göstergesi nöronal çığ, yani uzay-zamansal ölçek değişmeyen aktivite patlamaları, çalışma için sağlam bir yol. Çığ faaliyet uzun vadeli ölçümler için tam kontrollü koşullar altında düzlemsel bütünleşik çoklu-elektrod (MEA) ile sağlar kültürlü korteks yüzeysel katmanları geliştirme kendiliğinden ortaya çıkar.
Abstract
Korteks, herhangi bir giriş veya motor çıkış yokluğunda bile, kendiliğinden aktif. Gelişimi sırasında, bu faaliyet göç ve farklılaşma korteks hücre tipleri ve nöron bağlantıları 1 oluşumu için önemlidir. Olgun hayvan, sürekli aktivite uyarımlar sorunsuz bir şekilde gelecekteki eylemler hesaplamak için içine entegre edilmiş bir hayvan geçmiş ve mevcut durumunu yansıtır. Bu nedenle, devam eden yani spontan aktivite organizasyonu açık bir anlayış korteks fonksiyonu anlamak için bir ön koşuldur.
Çok sayıda kayıt teknikleri, korteks devam eden etkinlik, bireysel faaliyetleri geçici olarak ekstrasellüler Mikroelektronlar yerel alan potansiyeli (LFP) tespit edilebilir büyük olaylar toplamı, elektroensefalografi (EEG) magnetoencephalogram, (MEG birçok nöron oluşan olduğunu ortaya ) ve fonksiyonel manyetik rezonans görüntüleme (fMRI) BOLD sinyal. Mezoskopik ölçekte yüksek zamansal ve mekansal çözünürlükte (nöron binlerce) ile nöronal nüfus aktivite eğitim LFP şu anda tercih edilen bir yöntemdir. Ekstraselüler mikroelektrot, mekansal neighbored nöronlar mikrovolt yüzlerce LFP hızlı deplasmanlar sonucu faaliyetleri yerel olarak eşitlendi. Mikroelektronlar bir dizi kullanırken, bu tür sapmaların kuruluşların uygun uzay ve zaman içinde takip edilebilir.
Nöronal çığ ölçek değişmeyen 2,3 beyin devam eden nöronal aktivitenin uzaysal organizasyon açıklar . Anestezi sıçan 6 in vivo, in vitro 4,5 kurulan korteksin yüzeyel katmanlara özeldir ve uyanık maymun 7. Önemlisi, hem teorik ve ampirik çalışmalar 2,8-10 nöronal çığ, bilgi transferi ve bilgi işlem optimize korteks zarif dengeli bir kritik devletin dinamikleri işaret öneririz .
Tam kontrollü koşullar altında kararlı çığ aktivite kayıtları için izin nöronal çığ geliştirme, bakım ve düzenleme mekanizmalarını incelemek üzere, in vitro hazırlıklarında son derece yararlıdır . Geçerli protokol, düzlemsel, entegre mikroelektrot dizileri (MEA) da 11-14 yetişen Organotipik korteks kültürlerin, yani dilim kültürleri, yüzeysel katmanı gelişme yararlanarak in vitro nöron çığ nasıl çalışma açıklar .
Protocol
1. Uzun vadeli Kayıtlar için MEA ile Steril, Mühürlenebilir Cam Odası
- Güvenli ve sıkı bir kültür odası kapatılması için gerekli olan Teflon-plastik kap (as Cam), dişli Cam silindir alt parçacığı (Şekil 1A, B) (Aceglass) yaklaşık 2 mm kesilir. Su (3x) ve 200 kanıtı etil alkol içinde 5 dakika boyunca kaynar ile durulama Temizlik cam yüzük, kurumasını bekleyin.
- Kısım silikon çözümü MEA yüzey cam yüzük takmak için gereklidir. 15 ml parçaları karıştırın Sylgard 184 Silikon Elastomer Kit A & B iyice -20 ° C'de 1 ml alikotları mağazası, hava kabarcıklarını çıkarmak için 15 dakika bekletin
- MEA Tutkal cam halka (8x8 ızgara w / iç zemin elektrot, sıçan / fare için 30 mikron elektrot çapı 200/100 mikron elektrot arası mesafe) (Şekil 1A, B). 1 ml (23 ° C) silikon küçük iğne şırınga alın. ~ 60 ° C sıcak plaka üzerinde cilasız kesme cam yüzük, merkezi cam yüzük, MEA üzerinde güçlü bir mühür yüzük dışında etrafında ek bir silikon bir tabaka halinde yüzeye silikon uygulayın, 1 - 2 saat tedavi sağlar.
- 3x% 70 alkol ile takip deiyonize su ile durulayın bir laminer akış kaputu MEA odası ve kamara kapaklarını sterilize edin (3 x; son alkol içinde 10 dakika bekletin durulayın) UV ışığı odanın 10 dakika maruz kalma ve kapak iç takip . Otoklav MEA odası (120 ° C; ıslak, 45 dakika) ve kurumaya bırakın.
- Poli-D-lizin Coat MEA ile yüzey kültür odası içinde. Oldukça lipofilik yeni ÇÇA, elektrot ızgara çözüm tekrarlanan damlacık aspirasyon ile kaplamak. Kullanılan ÇÇA için çözümü ile odasının alt kapağı, aşırı sıvı aspire, laminer kaput içinde steril koşullar altında buharlaştırmak için izin verir. MEA kamara mühür depolama ve ileride kullanmak için kapağı takın.
2. Organotipik Kültürler Hazırlanması ve Büyüme İçin Gerekli Malzemeler
- Steril petri (Falcon, 100 x15 ~ 5 mm seviyesinde) içine dökün,% 0.9 NaCl içinde steril agar eritin, soğumaya bırakın ve depolama için Parafilm ile steril sargı. Katı agar kullanmak için 20 x 10 x 5 mm blokları kesin.
- Paketleme ile saklayın süper yapıştırıcı, örneğin Devcon Super Glue II, sterilite korumak için laminer akış kaputu içinde, önce% 70 EtOH sildi.
- % 50 D-Glukoz (SIGMA ultra, G7528), 40 gr glikoz 40 ml deiyonize kültür su (Sigma) ekleyerek hazırlayın. -20 ° C'de 2 ml alikotları saklayın
- 4 ml% 50 D-glukoz 500 ml Gey Dengeli Tuz çözüm ekleyin ve kullanmadan önce dondurucuda slush (sıvı / buz kristallerinin karışımı) soğuk.
- Yavaşça 10 dakika, girdap ve steril bir Petri kabı içine açık içerik decantate - 5 ml de-iyonize kültür su (kabarcıklarının oluşumunu önlemek, nazik sallamak) sulandırın tavuk plazma, çözüm 5 razı sağlar. Steril filtre (0.22 mikron gözenek filtresi; protein) -20 ° C'de plazma çözüm cryotubes içine kısım 350 ul (NuncTM), mağaza
- (NuncTM) kısım 40 ul cryotubes içine, (0.22 mikron gözenek filtre) buna göre, steril filtre sığır plazma sulandırın trombin, -20 ° C'de saklayın Çalışan bir çözüm için, Gey dengeli tuz solüsyonu w / D-Glikoz 375 ul trombin solüsyonunun 40 ul sulandırmak.
- At serumu, 200 ml Bazal Orta Kartal, 4 ml% 50 glikoz ve 200 mM L-glutamin 2 ml eklenir 100 ml Hank Tamponlanmış Tuzlu Çözüm 100 ml karıştırılarak 400 ml kültür ortamı hazırlayın. 8 hafta, 100 ml'lik PYREX şişeler içinde 4 ° C - 4 saklanabilir
- Mitoz inhibitörü 0.3 mM üridin, 0.3 mM ARA-C sitozin-β-D-arabinofuranoside ve 0.3 mM 5-floro-2'-dezoksiuridin, steril filtre, kısım 200 ul ve -20 ° C'de 6 mağaza karışımı hazırlayın - 12 ay.
3. Korteks ve ventral tegmental alan (VTA) Doku Diseksiyon (süre: <1hr)
- Prosedür verim korteks ve VTA ~ 12 sıçanlar ve fareler co-kültürler ve steril koşullar altında bir laminer akış kaputu içinde hazırlanan doku kesitlerinde. Doku toplanması için toplam süresi 1 saat daha edilmelidir.
- 1 - 2 postnatal gün (PND), sağlıklı, iyi beslenen yavrular (abdominal 'süt nokta' varlığı çöp boyutu ~ 10) alın. Burnu ile hafifçe bir yavru tutun, özgürce asmak için izin ve hızlı bir şekilde, keskin bir makas ile boyun üssünde başını kesmek.
- Beyin çıkarılması için, (iki yan makas keser) cilt kaldırmak ince göz makas (1 sagital orta hat kesme, 1 koronal korteks kes / serebellum kavşağı) ile açık kafatası kesmek. 4 kafatası flep geri çevirin. Bilenmiş bir spatula yardımı ile beyin, kaudal altında, koku ampul aracılığıyla cepheden önceden spatula kesmek. Kafatası beyin yavaşça kaldırın ve hızlı soğutma ve geçici depolama için steril, soğutulmuş, Gey çözümü slayt sağlar. 2 beyinleri için daha fazla 3.2 - 3.3 adımları tekrarlayın (toplam süresi: <20 dk).
- VTA almak içindoku, beyinlerinin küçük bir spatula kullanarak, kuru steril bir Petri kabı üzerine aktarın. Ayrıca yaklaşık 1 cm yanlara doğru hafifçe kaydırarak her bir beyin tarafından aşırı sıvı çıkarın. Beyin sapı, beyincik düzeyinde bir jilet kullanarak koronal, dikey kesim tarafından çıkarın.
- Dilimleme işlemi sırasında beyin mekanik stabilizasyon için disk montaj Tutkal agar bloğu. Superglue ince bir çizgi yerleştirin disk üzerinde agar bloğu önünde birkaç milimetre (agar tutkal dokunmaktan kaçının).
- Küçük bir spatula kullanarak, aşağı, frontal kutup aktarmak ve her beyin monte. Frontal kutuplu montaj disk yapıştırılmış olduğunu ve ventral taraf sırasında kesilmiş dilimleri kesme ve kolay asansör-off uygun mekanik stabilizasyon elde etmek için herhangi bir tutkal kalıntı bırakmadan agar temas ettiğinden emin olun
- Daldırın ve güvenli montaj diski bir vibratome tepsiye beyin montaj (örneğin Leica VT1000) dikkatlice steril, soğuk Gey çözümü ile dolu. Dikkatli bir şekilde temizlenmiş bir tıraş bıçağı ile (% 90 EtOH), en yüksek titreşim frekansı Ortabeyin koronal dilim kesilmiş ve nispeten düşük 400 500 mikron kalınlığında ileri hız. Emme ampul ters bir Pasteur pipeti kullanarak, transfer ve steril, soğuk Gey çözümü ile dolu 35 x 10 mm Petri kapları VTA içeren dilim toplamak (Şekil 1C; koronal plaka 18 - 15 E22 20).
- Korteks bölümleri için adımları tekrarlayın 3,2 - 3,6, ancak korteks ve serebellum arasındaki dikey kesme uygulamak ve frontal kutuplu forebrains kadar montaj. Striatum düzeyinde başlayarak yaklaşık 3 koronal dilim (350 mikron kalınlığında) gelecekteki korteks diseksiyon için toplanır.
- Kırık Jilet yapılan bir mikro-bıçak kullanarak, VTA (Şekil 1C) içeren bir stereomikroskop altında frontal korteks ve orta beyin alanları ~ 2 mm genişliğinde koronal bölümünde teşrih. Soğutulmuş Gey çözümü ile dolu küçük yemekler (örneğin odasının slaytlar) doku bölümleri ayrı ayrı toplayın.
4. Korteks ve VTA Doku Dilimleri Montaj MEA (Saat: <1 saat)
- Odak elektrot dizisi ile bir stereo mikroskop altında oda sıcaklığında pozisyon MEA. Merkezi temiz, tozsuz, 25 plazma ul damlacık ve steril elektrot dizi matris. Küçük spatulae kullanarak, plazma damlacık içine dikkatlice korteks ve VTA bölümüne kaydırın.
- MEA soğutma plakası üzerinde yerleştirin, görüntülemek ~ 15 s için izin soğuk plazma damlacık içine trombin 25 ul eklemek, sonra tekrar odaklayın. Trombin pipet kullanarak, MEA boyunca küçük dairesel hareketler ile plazma / trombin karışımı dikkatli bir şekilde yayılır. Kırılgan elektrot dizisi doğrudan temas etmeyin. Korteks, dizinin ikinci elektrot satır boyunca dorsal sınır dizi yavaşça yerleştirin. Bu şekilde, gelişmekte olan yüzeysel katmanları sonunda dizinin hatırlatma kapsayacaktır. VTA korteks bölümü (Şekil 1D), ventral sınır komşu yerleştirilir.
- Cap ve gevşek MEA / kültür montaj Plazma / trombin koagülasyon için izin kaputun içinde ~ 5 dakika boyunca oda sıcaklığında oturur yüksek nemi korumak için MEA odasına yakın. 3 kültürler için daha fazla 4.3 - Bu arada, 4.1 adımı tekrarlayın.
- 25 x 5 / 8 iğne ile 1 cc şırınga kullanarak kültür odasına küçük damlacıklar 600 ul kültür ortamı dikkatlice ekleyin.
- Sıkıca MEA odası ve yerde MEA / inkübatör (Şekil 1B) içinde sallanan depolama tepsisi kültür montaj kapatın. Için, 3 prosedürü hızlandırmak - 4 ÇÇA örtüşen dizileri monte edilebilir. Montaj süresi 12 ÇÇA <1hr olmalıdır.
- In vitro (DIV) 2 gün sonra, 10 ul mitoz inhibitörü ekleyin. Yenile kültür ortamı 4 SAYI az% 60 ve bundan sonra her 4 günde.
5. Elektrofizyolojik Kayıt ve Uyarıcı Üretimi
- MEA, her elektrot için 24 kHz ~ 10 dakika ~ 1 hafta 5,6 kayıt spontan aktivite sonra yerel alan potansiyeli (LFP) ve aksiyon potansiyelleri yangın nöron eğilimi önemli deplasmanlar arasındaki ilişki kurmak için (Donanım : MEA1060 w / kararma devre, x1200 kazanç, 12 bit A / D, aralık 0-4096 mV, Çok Kanallı Sistemleri Yazılım: MC_Rack). Zemin iç toprak elektrot yoluyla sağlanan, ya da dışarıdan bir Ag / AgCl yarım hücre ekleyerek.
- Ekstrasellüler başak aktivite (bant geçiren 300 - 200 Hz - 3.000 Hz) 1 bir bant geçiren filtre ile LFP ayırın. Spike aktivite daha off-line başak dizicili (örneğin Plexon A.Ş.) kullanarak tek ve çoklu ünite aktivitesi ayrılabilir. Hesapla başak her elektrot için ortalamaları tetikledi. Korteks kültürler için, ortalamalarının en çok tercih edilen kültür nöronal spike zaman olarak negatif LFP deplasmanlar (nLFP) belirleyecektir.
- (Şekil 2) -3 standart sapma (SD) LFP izleri gürültü her elektrot için bir eşik hesaplayınNLFPs pik süreleri ve genlikleri belirlemek çapraz eşiği (Şekil 2B, C). Ayrıntılar için: 2,4 görmek için; ve süresini ¨ t ardışık zaman aktif sanat (Şekil 2D aynı olan tüm elektrottan nLFPs birleştirerek dizi Spatiotemporal nLFP kümeleri tespit bir kez bin ¨ t (2 ila 8 ms ) seçti , 5).
- Nöronal çığ belirlemek için, her nLFP küme boyutunu hesaplamak, örneğin sayıda aktif elektrotlar veya nLFP amplitüdleri toplamı, bir boyut histogram oluşturmak ve çift-logaritmik koordinatlarda arsa. Nöronal çığ için, boyut dağılımı, düz bir çizgi ile çift logaritmik koordinatları 2 (Şekil 2E, F) yakınlaştıkları bir güç hukuk izler . Güç yasalar istatistiksel testler için 16 Bkz.
- Genlik G akım kontrollü uyaranlar (Stimulus jeneratör STG 1008, Çok Kanallı Sistemler) uygulanır bir elektrot seçerek doku uyarılmış tepkiler. - 200 uA genlik-S 50 μs genlik + S / 2 ve S ile 10 arasında 100 s: elektrot zararı azaltmak için, sınırlı bir dizi bipolar kare dalga ile tek şokların şarj nötr stimülasyon kullanabilirsiniz. Daha fazla bilgi için kullanım kılavuzu bakın.
- Dinamik aralık 9 stimülasyon sonra 500 ms sonra tüm elektrotlar 4 kHz örnekleme hızında LFP yanıtları kaydedildi kayıt uyaran kaydetmek için. Uyaran eserler azaltmak ve pre-amplifikatör doygunluğunu önlemek için uyarılması sırasında baş sahne yükselticiler keser devresi (Multi-kanal sistemleri) kararma kullanın.
6. Temsilcisi Sonuçlar:
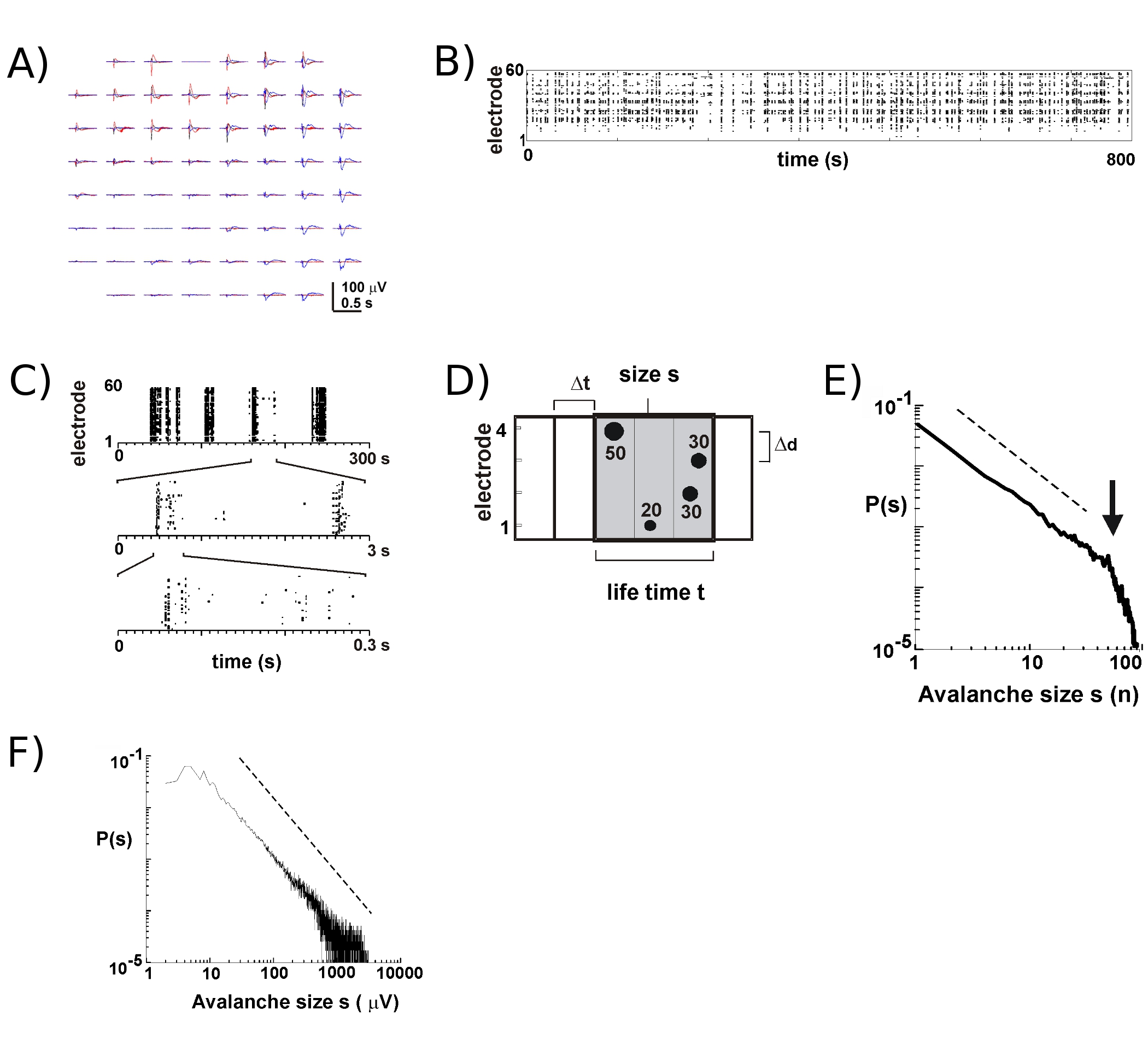
Yeni ÇÇA ile yaklaşık 8 - 9 - 10 kültürlerin birçok hafta boyunca ayakta kalacak. Uzun vadeli kayıtların çoğu bize birçok haftalık kurs 5 üzerinden bireysel kültürlerin gelişimini takip etmek sağlayan kültür ortamı, kuvöz içinde yer alır. Deneyler dayanarak, LFP kayıtları güvenilir 100'den fazla kültür günü için kullanılan ÇÇA ile elde edilebilir. Buna karşılık, ekstrasellüler başak aktivitesi daha güvenilir nispeten yeni ÇÇA (<40 kültür gün) ile ölçülür. Tipik bir deneyde, kültür odası mühürlü tutmak bağlı baş evre (Şekil 1B, sol) ile tepsiye depolama tepsisi (Şekil 1B, sağ) bir MEA aktarmak. 5 için korteks, korteks-VTA ortak kültürleri 6, anestezi sıçan 6 ve in vivo 7 uyanık maymun, yüzeysel katmanları nöronal çığ sırasında nöronal ateşleme de ağırlıklı olarak LFP pik negatif sapma yakın oluşur (nLFP .) Bu nedenle, yerel olarak senkronize nöronal grupların uzaysal organizasyon nLFPs dizi 17 uzay ve zaman içinde ortaya çıkması ölçerek tahmin edilebilir .
MEA Aktivite temporal kümeler halinde ortaya eğilimindedir, bu faaliyetin bir elektrot diğer sitelerde faaliyet eşlik ediyor. Birkaç saniye arayla meydana gelen 3 kümeleri komplo üzerine, bu tür faaliyet dönemlerinde LFP tipik dalga şekilleri Şekil 2A gösterilmiştir. Her küme için, negatif alan deplasmanlar 1 bir pencere içinde çeşitli elektrotlar görülebilir. S SD birden çok negatif bir eşik çapraz nLFP doruklarına açılan nLFP tepe kere şeklinde faaliyet nokta 'sütun' tesadüf nLFPs yakınında çeşitli elektrotlar (Şekil 2B) temsil eden bir raster uygun görüntülenmiştir. Bu faaliyetin Spatiotemporal organizasyonu oldukça karmaşık, daha az ya da çok düşük temporal çözünürlükte homojen görünüyor 'sütun', yüksek temporal çözünürlük ve böylece (Şekil 2C) ayrı kümeler oluşur. Aslında, Spatiotemporal nLFP kümeleri ortaya çıkması son derece kortikal ağlarda organize edilmektedir. Daha spesifik olarak, organizasyon nöronal çığ için ölçek değişmez. Bu, belirli bir zamansal çözünürlük ¨ t küme boyutları ihtimali hesaplanarak gösterilmiştir. Burada, kümeler aynı veya ardışık zaman kutuları (Şekil 2B) meydana nLFPs oluşmaktadır. Böyle bir küme boyutu Küme başına nLFPs veya Küme başına entegre nLFP amplitüdleri toplam sayısı olarak ifade edilir, küme boyutu dağılımları eğimi -1.5 2,4,5,7 gösterilmiştir güç hukuku, ortaya koymaktadır ( Şekil 2E, F). Bu dağıtım boyutları kxs, k sabit bir faktör oranı küme boyutları ölçek değişmeyen sipariş tanımlar unutmayın, s. bağımsız -1.5 k, Bu güç hukuku organizasyon dizi boyutu 2 bağımsız, temporal çözünürlük ¨ t 2 ve önemli nLFP tanımlamak için kullanılan eşik 7 fonksiyonudur. Nöronal grup büyüklüğü 7 ile nLFP genlik ölçekler, nLFPs ölçek değişmeyen örgütü ölçekli değişmeyen, yani fraktal, sipariş o yansıtırf yerel tüm boyutları içeren nöron gruplarının senkronize.

Şekil 1 (A) monte MEA dişli cam yüzük ile yan ve üst görünümleri ve buna karşılık gelen kap. (B) inkübatör içinde. Sol: headstage inkübatör koşul altında, tek bir kültür kayıt için izin monte. Sağ: kültür büyümesi için çok sayıda ÇÇA tutarak Tepsi. Yan jantlar: alternatif kültür gelişimi için gerekli olan batık ve atmosfere maruz kalan faz step motor kontrollü sallanan cihaz. (C), korteks VTA co-kültür için kullanılan koronal sıçan dilimleri için şematik çizimi. VTA ventral tegmental alan (VTA gri) içeren Cortex bölümlerde (sol) ve beynin bölümleri (orta, sağ), kırık çizgileri boyunca keserek elde edilir. Ctx: korteks; wm: beyaz cevher; cpu: striatum; VTA: pons: pontin alan. 15 ile ilgili koronal plakaları da 8, 18, ve 20 Bkz. (D), MEA ve kültürünün ilk 9 DIV gelişimi tek bir korteks-VTA ortak kültürler Yerleştirme ve büyüme. Kültür ve dizi ilerici genişleme, düzleşme unutmayın. Yansıtıcı doku parçaları, dejenere olmuş hücre ve doku artıkları göstermektedir. Sağlıklı doku, görünür ışık transillüminasyon altında opak ve grimsi.

Şekil 2 kortikal Organotipik kültürler Nöronal çığ. (A) Overplot dizi spontan aktivite üç dönem, birkaç saniye ile ayrılır. Her faaliyet dönemi dizi birçok elektrotlar üzerinde olumsuz LFP deplasmanlar (her renk bir faaliyet dönemine etiketler) oluşur unutmayın. (B) her elektrot nLFPs Negatif tepe kez bir aktivite raster içine monte edilir. 'Column' benzeri yapılar yakın senkron aktivite dönemleri göstermiştir. (C), yüksek bir zaman ölçeği senkronize görünür sütun yüksek zamansal ölçeklerde (3 zamansal ölçeklerde gösterilen) birden fazla sütun oluşur unutmayın. (D) nöronal çığ algoritması şematik gösterimi. Gürültü SD-x bir eşik geçen olumsuz LFP deplasmanlar (nLFP), 2 x 2 elektrot dizi zirve zaman ve genlik tanımlanır. NLFPs uzaysal organizasyon genişliği ¨ t gittikçe aktif zaman bidonları içine kümelenmiş. NLFP aktif site sayısı, yani elektrotlar da bir küme boyutu tarafından belirlenen (s = 4) veya nLFP amplitüdleri entegre toplamı (s = 130 μV). Yaşam süresi, ¨ t katları ile ölçülür. (E, F) Güç hukuk küme boyutu dağıtım nöronal çığ gibi kümeleri tanımlar. Dizi (burada 200 mikron) Δd belirli bir interelectrode mesafeler seçim dinamikleri dikkate alınmalıdır hangi belirli bir ¨ t tanıtır unutmayın. Daha spesifik olarak, hangi oran Δd / ¨ t güç hukuk eğimi α nöronal çığ 2,4,5 -1.5 yaklasik hangi ağ ortalama yayılma hızı yakındır. Lütfen Şekil 2'de büyük halini görmek için buraya tıklayın .
Discussion
1. Teknik konular:
- Steril ÇÇA hazırlanması ve kültür hazırlama tekniği. Steril koşullar altında bir laminer akış kaputu içinde yapılmaktadır. Nöronal aktivitenin etkileyen Antibiyotikler, hazırlanması ve kültür işlemi sırasında herhangi bir zamanda kullanılmaz.
- Plazma / trombin koagülasyon ve doku bağlılık MEA MEA Doku sağkalım üzerine plazma / trombin koagülasyon ve doku atmosfere maruz kalma süresi için gerekli zaman arasında dikkatli bir denge gerektirir. Atmosfere uzun süreli maruz kalma, doku dejenerasyonu tetikler kısa bir pıhtılaşma süresi, MEA kesitler erken dekolmanı riski vardır. Trombin çözüm gücü pıhtılaşma sürecinin hızını belirler, çünkü, sağlıklı kültürler MEA yüzeyine başarıyla takmak için çok önemli bir parametredir. Şu anda 1000 adet (1 NIH birim = 0.324 ± 0.073 mg 1KU) kullanarak en iyi sonuçları almak. Önemlisi, eksik kültür sırasında plazma rüptürlerinin teşvik mekansal heterojen koagülasyon Plazma / trombin solüsyonu sonuçları karıştırma. Bu plazma 'delik' kültür sağlığı ciddi etkiler ve Mikroelektronlar kültürünü ayırmak, böylece elektrofizyolojik kayıt kalitesinden ödün. MEA / doku montaj sırasında soğutma plakaları ile Çalışma koagülasyon yavaşlatır ve homojen bir karışım için plazma / trombin çözüm ve doku kesitlerinde doğru konumlandırma sağlar.
Benzer şekilde, tek damlacıkları orta, batığın koagülasyon sadece 5 dakika sonra kültür için kültür odası ekleyerek yüzey gerilimi nedeniyle doku müfrezelerin riskini büyük ölçüde azaltır. Başarılı bir kültür MEA düzleştirmek ve biraz eksik doku MEA kişiyi ya da doku dejenerasyonu (örneğin Şekil 1D) herhangi önemli belirtileri olmaksızın, hafta boyunca sağlıklı bir büyüme gösteren genişletmek olacaktır. - Doku diseksiyonu. Micro bıçak diseksiyonu süreci büyük ölçüde iyileştirilmiştir. Geniş ~ 2 mm 'bıçaklar' pense kullanarak bölmek ve bir neşter sahibi ile onları tutun, çift kenar Jilet (100.050-00 - kırılabilir bisturi ağızları Güzel Bilim Araçları) kullanın. Bıçağın düz, dikey bir firma hareket kullanarak koronal dilim disseke doku örnekleri böylece büyük ölçüde geliştirerek genel kültür sağlık doku çekerek nedeniyle mekanik stresi azaltarak.
- Doku soğutma. Hazırlanması sırasında dilimleri ve doku bölümleri uygun soğutma kültür başarısı için şarttır. Biz bir metal disk altına iliştirilmiş peltier elementleri inşa ısmarlama soğuk tabak kullanın. Peltier elemanı tarafından üretilen ısı, soğuk su perfüzyon ile kaldırılır. Bu hazırlık süresi büyük ölçüde azaltır ve hazırlık her aşamasında soğutma standartlaştıran (Dold Laboratuvar ve Mühendislik 131 Plantation Dr Seguin, TX 78.155; (830) 560-1471)).
- İnkübatör durumu hassas iç sallanan koşulları ile ısmarlama bir inkübatör ÇÇA dilim yetiştirilmesi başarımız için çok önemli olmuştur . (Şu anda piyasada bulunan) Multichannelsystems tarafından in-house tasarım orijinal dayanarak, iç sallanan cihaz iki büyük yan tekerleklere bağlı tepsiler oluşur. Step motorlar ve bilgisayarlı kontroller (, açı sallanan hız sallanan ve aralıklı duraklar) kesin bir sallanan yörünge için izin verir. Sonuçta, dilim kültürleri yavaş değişmesinde atmosfer ve kültür ortamına maruz kalması gerekir. Geleneksel yaklaşım, orada uzun ekseni boyunca yavaş yavaş döndürmek dar tüpler içine dilim kültürler yerleştirmektir. Burada, yavaş rotasyon nedeniyle mekanik stres dönme kendisi üretemez, ve dönme hızı, yaklaşık 5 optimum 'besleme / nefes döngüsü elde etmek için yeterince yüksek - 10 dakika süresi. MEA odası, toplam hacmi ~ 2 cc, ve doku kendisi tarafından orta klima için gerekli küçük bir kültür orta hacimli daha kompakt iç, önemli bir sorun teşkil etmektedir. ± ~ 70 ° açı (çevrim süresi: ~ 200 s) arasında MEA sallanan, sıvı faz ve atmosfer arasındaki kültür geçişleri gibi sallanan hızı yavaşlatmadan, ve ekstrem açılar atmosfere uzun pozlama için sallanan durdurulması kültür yaşam için gereklidir.
2. Nöronal çığ çalışma korteks kültürler Gelişimsel yaş
Akut sıçan korteks dilim yaygın PND 0 - 1 ve MEA birçok hafta kültür. Erken çalışmalar tek korteks dilim kültürleri, in vitro birkaç hafta sonra, in vivo hücre sınıfları 18,18-21 kolayca mukayese edilebilir, tanımlanabilir hücre tipleri ile katmanlı yapısını korumak olduğunu açıkça ortaya koymuştur. Bu in vitro sistemde katmanlı organizasyon uygun talamik çalışma için kullanılır olmuştur22-24 geliştirme sırasında korteksin innervasyon yanı sıra, striatum 25,26 olarak subkortikal yapıları sürüş için. Aslında, beyin bölgeleri içinde ve arasında nöronal bağlantıları oluşumunda özgüllüğü örneğin korteks-bazal ganglia devresi 27-30, kompleksin yapımı için çok detaylı bir projeksiyon sistemleri yeniden ele in vitro sistemlerde sağlar.
- 4 in vitro, tek korteks dilim 31 ve korteks dilim, striatum 26 veya talamus spontan 32 gösteri ile birlikte kültürlü, yukarı ve aşağı genellikle üretan anestezisi sıçan 33 in vivo olarak bulundu devletlerin 6 hafta sonra -Devletlerin bu kadar ince zamansal organizasyon damgasını taşıyan iç içe θ ve γ-salınımlar piramidal nöronlar ve hızlı spike GABAerjik internöronlar 31 elektrofizyolojik olgun bir ağ göstergesidir. Daha da önemlisi, dopamin D2 reseptör stimülasyonu yokluğunda, parvalbumin pozitif kortikal internöronlar olgunlaşması korteks dilim kültürler 34 yaklaşık 2 hafta gecikti. Bu bulgular doğrultusunda, gelişim süresi içerisinde yuvalanmış θ, β ve γ-salınımlar in vivo korteks dilim için projelendirme dopaminerjik nöron içeren ventral tegmental alan (VTA) ile birlikte kültüre zaman buna uygun korteks 6.
Bu çalışmaların en önemlisi olgun hızlı GABAerjik inhibisyonuna bağlıdır ve korteks 2,4 yüzeysel katmanları bulunan nöronal çığ, eğitim, büyük bir özenle kortikal doku olgunlaşmasını sağlamak için alınması gerektiğini göstermektedir. Nöronal çığ 2 saat boyunca tek korteks kültürlerde ortaya çıkan, in vivo gelişimine uygun bir gelişim süresi içerisinde gerektiren 5 hafta 4, korteks ortak kültür korteks dilim dilim uygun bir dopamin reseptör stimülasyonu, örneğin gerekir VTA 6 ile.
Disclosures
Çıkar çatışması ilan etti.
Acknowledgments
Bu çalışma, Ulusal Ruh Sağlığı Enstitüsü, Ulusal Sağlık Enstitüleri İntramural Araştırma Programı Bölümü (DIRP) tarafından finanse edildi.
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}