

Summary
Надежный способ для изучения нейронных лавины, то есть масштабно-инвариантной пространственно-временные всплески активности, что свидетельствует о критической динамики состояния в коре головного мозга. Лавины возникают спонтанно в развивающихся поверхностные слои культурной коры, который позволяет для долгосрочных измерений деятельности с плоскими интегрированной многоэлектродной массивы (МЭС) в точно контролируемых условиях.
Abstract
Коры спонтанно активны, даже при отсутствии какого-либо конкретного входа или мощность двигателя. Во время разработки, эта деятельность имеет важное значение для миграции и дифференциации типов коры клеток и образованию нейронные связи 1. В взрослого животного, текущей деятельности отражает прошлое и настоящее состояние животного, в котором сенсорные стимулы интегрированы для расчета будущих действий. Таким образом, четкое понимание организации работы, проводимой т.е. спонтанную активность является необходимым условием для понимания коры функции.
Многочисленные методы записи показали, что постоянная деятельность в коре головного мозга состоит из множества нейронов, отдельных видов деятельности временно сумму в более крупные события, которые могут быть обнаружены в местных потенциальном поле (LFP) с внеклеточных микроэлектродов, или в электроэнцефалограмме (ЭЭГ), magnetoencephalogram (МЭГ ), а BOLD сигнал с функциональной магнитно-резонансной томографии (МРТ). LFP в настоящее время является методом выбора при изучении нейронной активности населения с высоким временным и пространственным разрешением в масштабе мезоскопических (несколько тысяч нейронов). На внеклеточных микроэлектродов, локально синхронизированы деятельности пространственно Соседями нейронов приводит к быстрому отклонений в LFP до нескольких сотен микровольт. При использовании массива микроэлектродов, организаций таких отклонений может быть удобно контролировать в пространстве и времени.
Нейронные лавины описывать масштабно-инвариантной пространственно-временной организации работы, проводимой активности нейронов в мозге 2,3. Они являются специфическими для поверхностных слоев коры, установленных в пробирке 4,5, в естественных условиях в анестезии крыс 6, а в бодрствует обезьяны 7. Важно отметить, что теоретические и эмпирические исследования показывают, что 2,8-10 нейронов лавин указывают изысканно сбалансированной критической динамики состояния коры, которая оптимизирует передачу информации и обработки информации.
С целью изучения механизмов нейронных развития лавины, технического обслуживания и регулирования, в пробирке препараты весьма полезным, поскольку они позволяют для стабильной записи лавинной активности в точно контролируемых условиях. Текущий протокол описывает, как изучение нейронных лавин в пробирке, пользуясь поверхностным слоем в развитии органотипической культур коры, то есть срез культуры, выращенные на плоской, интегрированные массивы микроэлектрода (MEA, см. также 11-14).
Protocol
1. Стерильные, Уплотненная.Служит палаты Стекло с МЭС для долгосрочной записи
- Резьбовые цилиндров стекла с тефлоновым пластиковый колпачок (Ace Glass), необходимых для безопасного и плотно закрытия камеры культуры, режутся (Aceglass) примерно 2 мм от нижней нити (рис. 1а, б). Чистая кольца стекло, промыв водой (3 раза) и кипячения в течение 5 мин в 200 доказательство этилового спирта, дайте высохнуть.
- Решение Алиготе кремния необходимо приложить стекла кольца к поверхности МПС. Смешайте 15 мл частей и Б Sylgard 184 Kit Эластомер Силиконовые тщательно, дайте постоять 15 мин для удаления пузырьков воздуха, храните в 1 мл аликвоты при -20 ° C.
- Клей стеклянное кольцо для MEA (сетка 8x8 Вт / внутреннего электрода землю, 30 мкм электродов диаметром 200/100 мкм межэлектродного расстояния для крысы / мыши) (рис. 1а, б). Возьмите 1 мл кремния (23 ° С) в шприц с иглой небольшое. Нанесите силиконовый для неполированный поверхность среза стеклянное кольцо, в центре стекла кольцо на МЭС, нанесите дополнительный слой кремния вокруг внешней стороны кольца для более сильного уплотнения, не говоря вылечить в течение 1 - 2 часа при температуре ~ 60 ° C на горячую тарелку.
- Стерилизовать МЭС камеры и камеры шапки в ламинарном потоке 3x промыть в деионизированной воды, а затем на 70% спирте (3 х; для последнего полоскания пусть сидят в течение 10 мин в спирте), затем 10 мин экспозиции камеры и крышки интерьера к ультрафиолетовому излучению . Автоклав МЭС камеры (120 ° С; мокрые, 45 мин) и дайте высохнуть.
- Пальто МЭС поверхность внутри культуры камеры с поли-D-лизина. Для новых МПС, которые являются довольно липофильных, пальто повторными капли стремление решение от электрода сетки. Для подержанных МПС, накрыть дно камеры с раствором, аспирация лишней жидкости, позволяют испаряться в стерильных условиях в ламинарном капотом. Прикрепить заглушки МЭС камеры для хранения и дальнейшего использования.
2. Ингредиенты, необходимые для подготовки и рост органотипической культур
- Растворите стерильным агаром в 0,9% NaCl, разлить в стерильные чашки Петри (Falcon, 100 x15; ~ 5 мм уровне), дать остыть, и стерильные обертывание с парафильмом для хранения. Вырезать 20 х 10 х 5 мм, блоки из твердого агара для использования.
- Магазин супер клей, например, Devcon супер клей II, с упаковкой протирать с 70% этанола, прежде чем открыть, внутри ламинарном боксе сохранить стерильность.
- Подготовка 50% D-глюкозы (SIGMA ультра, G7528), добавив 40 г глюкозы в 40 мл деионизированной воды культура (Sigma). Хранить в 2 мл аликвоты при -20 ° C.
- Добавьте 4 мл 50%-D-глюкозы до 500 мл сбалансированного солевого раствора Гей, и холод в слякоть (смесь жидких / кристаллы льда) в морозильной камере перед использованием.
- Развести куриные плазме в 5 мл деионизированной водой культурой (взболтайте нежная, следует избегать образования пузырьков), пусть решение согласиться на 5 - 10 мин, мягко вихрем, и декантата ясно содержание в стерильную чашку Петри. Стерильный фильтр (0,22 мкм поры фильтра; белка) плазмы решение, аликвоту 350 мкл в cryotubes (NuncTM), хранить при -20 ° C.
- Развести тромбина из бычьей плазмы соответственно, стерильный фильтр (0,22 мкм поры фильтра), аликвоту 40 мкл в cryotubes (NuncTM), хранить при -20 ° C. Для рабочего раствора, развести 40 мл раствора тромбина в 375 мкл сбалансированный солевой раствор Гей в ж / D-глюкозы.
- Подготовка 400 мл питательной среды путем смешивания 100 мл лошадиной сыворотки, 200 мл базальной среды Eagle, 100 мл буферизацией Хэнка солевой раствор в которых 4 мл 50% глюкозы и 2 мл 200 мМ L-глутамина добавлены. Может храниться 4 - 8 недель в 100 мл PYREX бутылки при температуре 4 ° C.
- Подготовка митоза ингибитор путем смешивания 0,3 мМ уридина, 0,3 мм ARA-C цитозин-β-D-арабинофуранозида, и 0,3 мм 5-фтор-2'-дезоксиуридина, стерильный фильтр, аликвоту 200 мкл и хранить при температуре -20 ° С в течение 6 - 12 месяцев.
3. Cortex и вентральной области покрышки (VTA) тканей Dissection (время: <1 час)
- Процедура дает коры и VTA срезах тканей для ~ 12 со-культур от крыс и мышей, и готова в ламинарном боксе в стерильных условиях. Общее время для ткани коллекции должны быть чем на 1 час.
- Возьмите здоровой, хорошо питается щенков (помет размер ~ 10; наличие брюшной 'молоко пятно »), на 1 - 2 день после рождения (PND). Держите щенка осторожно морду, позвольте ему висеть свободно, и быстро обезглавить у основания шеи с острыми ножницами.
- Для удаления мозга, удалить кожу (два боковых ножницами разрезы), вырезать череп с мелкой ножницеобразный глаз (1 сагиттальной срединной линии разреза, 1 корональные вырезать на коре / мозжечка перехода). Отразить обратно все 4 клапанами черепа. С заостренными шпателем, вырезать фронтально через обонятельные луковицы, заранее шпателем каудально под мозга. Аккуратно поднимите мозг из черепа и дайте ему скользить в стерильные, охлажденные, решение Гей для быстрого охлаждения и временного хранения. Повторите шаги 3,2 до 3,3 на 2 больше мозгов (общее время: <20 мин).
- Для получения VTAткани, передачу мозги на стерильные, сухой чашке Петри с помощью небольшого шпателя. Далее удалите лишнюю жидкость, осторожно скользящего каждый мозг около 1 см в сторону. Удалить ствола мозга по корональной, вертикальным разрезом на уровне мозжечка использованием лезвия бритвы.
- Клей агар блока на диске для установки механической стабилизацией мозг во время нарезки процедуры. Место тонкая линия суперклея несколько миллиметров перед агар блока на диске (во избежание клей трогательные агар).
- С помощью небольшой лопаточки, передачи и горе каждого мозга, фронтальные полюса вниз. Убедитесь, что фронтальные полюса приклеены к установке диска и вентральной сторон сенсорного агар без каких-либо остатки клея для того, чтобы достичь правильной механической стабилизации во время резки и легко старт резания ломтиками.
- Осторожно погрузите и надежного крепления диска с мозгом сборки в лоток vibratome (например, Leica VT1000), заполненной стерильным, охлажденные решение Гей это. С тщательно очистить лезвие бритвы (90% этанола), нарезать ломтиками корональных среднего мозга на самом высоком частоты колебаний и относительно низкой скорости движения при толщине 400 500 мкм. Использование перевернутого пипетку Пастера с всасывающим лампочка, передавать и собирать ломтиками содержащие ВТА в 35 х 10 мм чашки Петри, заполненной стерильным, охлажденные решение Гей (рис. 1С, см. также корональные пластина 18 - 20 на E22 в 15).
- Для коры разделах, повторите шаги 3.2 - 3.6, но применяют вертикальный разрез между корой и мозжечком, и монтировать forebrains с фронтальной полюсом вверх. Около 3 корональных ломтиками (толщиной 350 мкм), начиная с уровня полосатого тела собираются в будущем рассечение мозга.
- Использование микро-нож из сломанной лезвия бритвы, рассекать ~ 2 мм шириной корональной части лобной коре мозга и районов, содержащих VTA (рис. 1в) под стереомикроскопа. Сбор срезах тканей отдельно в небольших блюд (например, слайды камеры), заполненные раствором охлажденного Гей это.
4. Монтаж Cortex и фрагменты тканей VTA на MEA (Time: <1 час)
- Позиция МПС при комнатной температуре в стерео микроскоп с электродом массив в центре внимания. Центр 25 мкл капли плазмы на чистой, без пыли, и стерильные матрицей массив электродов. Использование малых spatulae, аккуратно вставьте коры и VTA раздел в плазму капли.
- Место МЭС при охлаждении пластины, переориентировать зрения, не говоря охладить в течение ~ 15 с, затем добавляют 25 мкл тромбина в капле плазмы. Использование кончиком пипетки тромбина, тщательно распространения плазмы / тромбина смеси с небольшими круговыми движениями по всему МПС. Не прикасайтесь к хрупкому массив электродов непосредственно. Аккуратно позиции коры на массив с спинной границы по второй ряд электрод массив. Таким образом, развивающиеся поверхностные слои, в конечном счете покрытие напоминание о массиве. VTA находится рядом с вентральной границе раздела кора головного мозга (рис. 1D).
- Кап и свободно закрыть камеру МЭС сохранить высокую влажность в то время как МЭА / культура сборки сидит за ~ 5 мин внутри капота при комнатной температуре, чтобы для плазменных / тромбина коагуляции. Между тем, повторите шаг 4,1 - 4,3 на 3 больше культур.
- Аккуратно добавить 600 мкл культуральной среды в малых капель культуры камеры использованием 1 мл шприца с 25 х 5 / 8 иглы.
- Плотно закрыть МЭС камеру и место МЭС / культура сборки на качалке лоток хранения внутри инкубатора (рис. 1В). Чтобы ускорить процедуру, 3 - 4 МПС могут быть собраны в перекрывающихся последовательностей. Ассамблея времени в течение 12 МПС должно быть <1 час.
- Через 2 дня в пробирке (DIV), добавить 10 мкл митоза ингибитора. Обновить культуру средств массовой информации на 60% за 4 DIV и каждые 4 дня после этого.
5. Электрофизиологические Запись и Стимул поколения
- Чтобы установить связь между значительными отклонениями в местных потенциальном поле (LFP) и тенденция нейронов уволить потенциалов действия, после ~ 1 недели 5,6 запись спонтанной активности при 24 кГц ~ 10 мин от каждого электрода MEA (Hardware : MEA1060 ж / гашения цепи, x1200 усиления, 12-битный A / D, диапазон 0-4096 мВ, многоканальных системах; Программное обеспечение: MC_Rack). Земля предоставляется либо через внутренний электрод землю, или внешне, добавив Ag / AgCl половину клетки.
- Отдельные LFP с полосовой фильтр 1 - 200 Гц от внеклеточных всплеска активности (полосы пропускания 300 - 3000 Гц). Спайк деятельности могут быть разделены на одно-и многоквартирных деятельности с использованием автономной шип сортировщики (например Plexon Inc.) Рассчитать шип срабатывает средние для каждого электрода. Для культур коры, большинство средних будет выявлять отрицательные отклонения LFP (nLFP) как предпочтительное время нейронных пики в культуре.
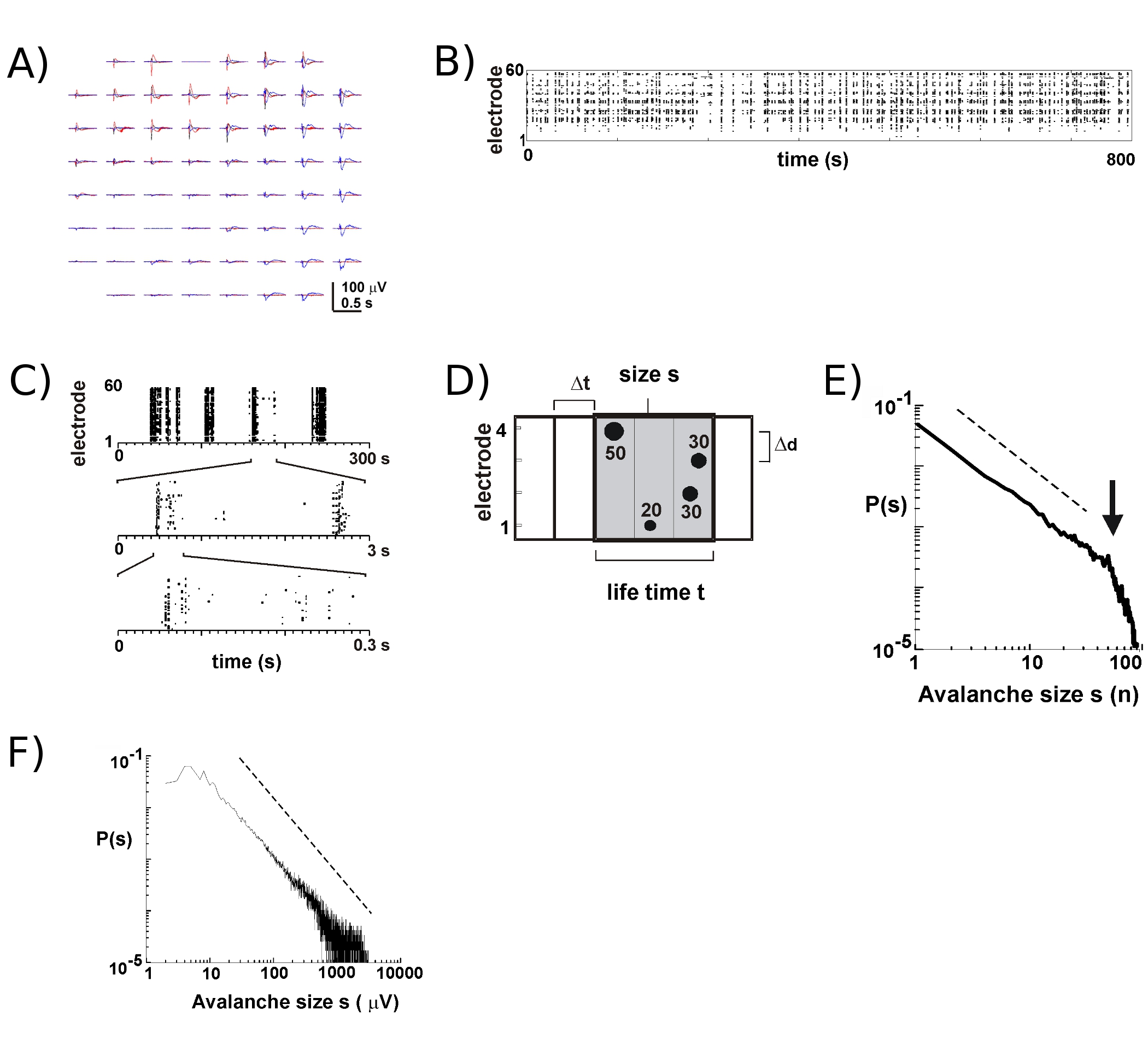
- Рассчитать для каждого электрода порог -3 стандартных отклонений от шума (SD) с LFP следов (рис. 2), Определить пиковые периоды и амплитуды nLFPs, которые пересекают порог (рис. 2, б, С). Выбрал время бен Δt (например, между 2 - 8 мс) и определить пространственно-временные кластеры nLFP на массиве путем объединения nLFPs от всех электродов, которые находятся в той же последовательных побегов времени длины Δt (рис. 2D; подробнее см. 2,4 , 5).
- В целях выявления нейронов лавин, рассчитать размер каждого кластера nLFP, например, число активных электродов или сумма nLFP амплитуд, построить размер гистограммы, и сюжет в двойных логарифмических координатах. Для нейронов лавин, распределение по размерам следует степенному закону аппроксимируется прямой в двойных логарифмических координатах 2 (рис. 2E, F). Смотрите 16 для статистических тестов на власть законов.
- Выявить вызвала отклики в ткани, выбрав электрод, через который управляемый током стимулов с амплитудой S применяются (Стимул генератор STG 1008, многоканальных системах). Для уменьшения повреждений электродов, использование диапазона ограничено, заряда нейтральными стимуляция одного потрясения с биполярным квадратных сигнала: 50 мкс с амплитудой-S со 100 мкс с амплитудой + S / 2 и S от 10 - 200 мкА. См. руководство пользователя для получения дополнительной информации.
- Для записи динамический диапазон 9, запись стимул записал LFP ответы на 4 кГц частотой дискретизации на всех электродах следующие 500 мс после стимуляции. Использование схемы гашения (Multi-канальные системы), который отключает усилитель головы сцене во время стимуляции, чтобы уменьшить стимул артефактов и, чтобы предотвратить предварительный усилитель насыщения.
6. Представитель Результаты:
С новыми МЭС около 8 - 9 из 10 культур выживет в течение многих недель. Большинство наших долгосрочных записей происходят внутри инкубатора в культуральной среде, что позволяет нам следить за развитием отдельных культур в течение многих недель 5. Исходя из наших экспериментов, LFP записи могут быть надежно, полученные с МПС использован для более чем за 100 дней культуры. В отличие от внеклеточных всплеска активности более надежно измерена с относительно новыми МПС (<40 дней культуры). В типичном эксперименте, мы переносим МЭС от хранения лоток (рис. 1В, справа), чтобы лоток с головы этапе прилагается (рис. 1В, слева) поддержания культуры камеры опечатаны. Для коры 5, коре-VTA совместно культур 6, а также в анестезии крыс 6 и бодрствования обезьян в естественных условиях 7, нейронов в нейронной лавин в поверхностных слоях происходит преимущественно близко к пику отрицательное отклонение LFP (nLFP ). Таким образом, пространственно-временной организации местного синхронизированных нейронов группы можно оценить, измеряя возникновения nLFPs в пространстве и времени на массиве 17.
Деятельность по МЭС имеет тенденцию возникать в височной кластеров, например, что активность на одном электроде сопровождается деятельность на другие сайты. Типичные осциллограммы LFP в такие периоды деятельности показаны на 2A фигура, более чем на 3 построение кластеров происходит несколько секунд. Для каждого кластера, отрицательные отклонения поле можно увидеть на нескольких электродов в окне 1 с При извлечении nLFP пики, которые пересекают порог нескольких негативов SD, деятельность в форме nLFP часы пик удобно визуализировать растровые, в котором "столбцов" точек представляют около совпадающих nLFPs на различных электродов (рис. 2б). Пространственно-временной организации этой деятельности является довольно сложным; 'столбцов, которые кажутся более или менее однородные по низким временным разрешением, состоят из отдельных кластеров на более высоком временном разрешении и т.д. (рис. 2). На самом деле, появление пространственно-временных кластеров nLFP имеет высокую степень организации в кортикальных сетей. В частности, организация масштабно-инвариантной для нейронов лавин. Об этом свидетельствуют вычисления вероятности размеров кластеров в данный временной Δt разрешения. Здесь, кластеры состоят из nLFPs, которые происходят в том же или раз подряд бункеров (рис. 2D). Когда размер такого кластера выражается в общей численности nLFPs в кластере или интегрированные nLFP амплитуд на кластер, размер кластера распределения показывает, степенному закону, наклон которой было показано, что -1,5 2,4,5,7 ( Рис. 2E, F). Обратите внимание, что это распределение определяет масштабно-инвариантной упорядочении размеров кластеров, что это соотношение размеров с до kxs, где к постоянного множителя, равен к -1,5, которая не зависит от s. Эта организация степенному закону не зависит от размера массива 2, временное разрешение Δt 2, и порог используется для выявления значительных отклонений nLFP 7. Потому что nLFP амплитуды весы с нейронной размер группы 7, масштабно-инвариантной организации nLFPs отражает масштабно-инвариантной, то есть фрактал, о заказеF локально синхронизированных нейронов группы, которые включают в себя все размеры.

Рисунок 1. () Сбоку и сверху вид МЭС с резьбовым кольцом стекла установлены и соответствующие крышки. (B) Внутренний вид инкубатора. Слева: headstage крепление позволяет для записи с одной культуры при условии инкубатора. Справа: лоток проведение многочисленных МПС для роста культуры. Сторона колеса: шаговый двигатель контролируемой качалки устройство для переменного погружения и атмосферы, подвергшихся воздействию фазу, необходимых для роста культуры. (C) Схематическое изображение для корональных ломтиками крысы используется для Cortex-VTA совместно культур. Cortex разделов (слева) и среднем мозге разделы (в центре справа), содержащие VTA вентральной области покрышки (VTA, серые) получаются разрезанием вдоль ломаных линий. CTX: кора головного мозга; WM: белое вещество; Процессор: полосатом теле; ВТА моста: мостовой области. См. также соответствующие корональных пластин 8, 18 и 20 на 15. (D) размещения и рост коры одного-VTA совместно культур на рубеже тысячелетий и ее развитие в течение первых 9 DIV в культуре. Обратите внимание, уплощение культуры и ее постепенное расширение на массив. Отражающие частей ткани указывают выродились клеток и тканей мусора. Здоровые ткани является непрозрачным и серовато под просвечивание видимым светом.

Рисунок 2. Нейронные лавин в корковых органотипической культур. (А) Overplot трех период спонтанной активности на массив, разделенные на несколько секунд. Обратите внимание, что каждый вид деятельности периода состоит из отрицательных отклонений LFP на многих электродов на массив (каждый цвет этикетки одного вида деятельности периода). (Б) Отрицательные раз пик nLFPs от каждого электрода собраны в растровых деятельности. "Column'-подобные структуры указанных периодов около синхронной активности. (C) Обратите внимание, что столбцы, которые появляются высоко синхронизирован на одном временном масштабе состоит из нескольких столбцов на более высоких временных масштабов (3 временных масштабах показаны). (D) Схематическое изображение нейрона алгоритм лавину. На 2 х 2 множество электрода время пика и амплитуды отрицательных отклонений LFP (nLFP) пересечения порога-х УР шума выявлены. Пространственно-временная организация nLFPs кластеризован в последовательно активное время закромах шириной Δt. Размер кластера определяется как количество активных сайтов, т.е. электроды с nLFP (S = 4), либо интегрированы сумму nLFP амплитуды (а = 130 мкВ). Срок службы измеряется в кратных Δt. (E, F) Питание права в кластере определяет распределение по размерам кластеров нейронов лавин. Обратите внимание, что выбор того или иного расстояния межэлектродном Δd для массива (в данном случае 200 мкм) вводит особый Δt, при которой динамики должны быть соблюдены. В частности, отношение, в котором Δd / Δt приближает среднюю скорость распространения в сети, при которой наклон α власти закона приблизительно -1,5 для нейронов лавин 2,4,5. Пожалуйста, нажмите здесь , чтобы видеть большую версию рисунке 2.
Discussion
1. Технические вопросы:
- Стерильная техника. Подготовки МПС и культуру приготовления всех выполненных в ламинарном боксе в стерильных условиях. Антибиотики, которые влияют на активность нейронов, которые не используются в любое время в ходе процесса подготовки и культуры.
- Плазма / тромбина коагуляции и ткани присоединении, в МПС. Ткани выживания на МЭА требует тщательного баланса между время, необходимое для плазмы / тромбина коагуляции и время ткань воздействию атмосферы. Короткое время свертывания риски преждевременной отслойке секции из МЭС, в то время длительного пребывания в атмосферу вызывает дегенерацию тканей. Потому что силы раствора тромбина определяет скорость процесса коагуляции, это очень важный параметр для успешного присоединения, здоровых культур на поверхности МПС. Получим лучших результатов, используя 1000 единиц (1KU, 1 единица = NIH 0,324 ± 0,073 мг). Важно отметить, что неполное перемешивание плазмы / тромбина решение приводит к пространственно неоднородной коагуляции содействия плазменных разрывов во время культивирования. "Дыры" Эти плазменные серьезно повлиять на культуру здоровья и отделить культуру от микроэлектродов, тем самым ставя под угрозу качество электрофизиологические записи. Работа с охлаждающих пластин во время МЭА / ткани сборки замедляет коагуляцию и обеспечивает однородное перемешивание плазмы / тромбина решение и правильное позиционирование срезах тканей.
Аналогичным образом, путем добавления среды в одной капли, чтобы культура камеру, чтобы погрузить культуре уже через 5 мин коагуляции, что значительно снижает риск ткани отряды из-за поверхностного натяжения. Успешной культуры будет сглаживаться по МЭС и немного расширить показывает здоровый рост в течение недель, без каких-либо серьезных признаков неполного ткани МЭС контакт или ткани дегенерации (например, рис. 1D). - Ткань рассечение. Micro ножи значительно улучшили нашу вскрытия процесса. Мы используем двойное ребро лезвия (Fine инструменты Наука - хрупкие скальпель лопастей - 100050-00), из которой мы расстались ~ 2 мм "лезвия", используя плоскогубцы, и держать их с помощью скальпеля держателя. Образцы тканей расчлененное из корональной слайс, используя гладкие, вертикальные движения фирмы лезвие тем самым значительно уменьшая механическое напряжение из-за вытягивания ткани общего повышения культуры здоровья.
- Ткань охлаждения. Правильное охлаждение ломтиками и срезах тканей во время подготовки имеет важное значение для культуры успех. Мы используем заказ холодных блюд построены из элементов Пельтье прилагается внизу, чтобы металлический диск. Тепла от элемента Пельтье удаляется через холодную воду перфузии. Это значительно сокращает время на подготовку и стандартизирует охлаждения в каждой стадии подготовки (Дольд Labs и инженерно 131 Плантации доктор Seguin, TX 78155;. (830) 560-1471)).
- Инкубатор состоянии. Заказных инкубаторе с точными внутренними условиями качалка имеет решающее значение для нашего успеха в выращивании ломтики на МПС. На основе оригинального внутреннего дизайна Multichannelsystems (в настоящее время не доступен коммерчески), внутреннее устройство качания состоит из лотков, которые прикреплены к двум большим колесам сторону. Шаговые двигатели и автоматизированные средства управления позволяют для точной траектории качания (угол качания, покачиваясь скорости и прерывистый пауз). В конце концов, ломтик культур должны быть подвержены атмосферы и культуральной среде в медленном чередовании. Традиционный подход заключается в размещении срез культур в узкие трубы, которые медленно вращаться вместе там длинная ось. Здесь, медленное вращение не производит механическое напряжение из-за вращения себя, а скорость вращения достаточно высока, чтобы получить оптимальную «кормления / дыхание" цикла примерно на 5 - 10 мин продолжительность. Более компактный интерьер МЭС камере, его общий объем ~ 2 см, а также небольшой объем питательной среды для среды кондиционирования от самой ткани, представляет собой значительную проблему. По качалка МЭА в пределах ± ~ 70 градусов (время цикла: ~ 200 с), замедление скорости качания, как культура переходы между жидкой фазы и атмосферу, и остановить качание на крайние углы в течение длительного контакта с атмосферой огромное значение для культуры выживания.
2. Развивающие возраста культур коры для изучения нейронных лавин
Острые ломтики из коры крысы обычно принято на PND 0 - 1 и культивировали в течение многих недель по МЭС. Ранние исследования ясно показывают, что одной культуры срез коры, после нескольких недель в лабораторных условиях, поддерживать слоистую структуру с конкретными типами клеток, которые можно легко по сравнению с в естественных классов ячейки 18,18-21. Слоистой организацией в этой системе в пробирке была удобно использовать для изучения таламусаиннервации мозга при развитии 22-24, а также за вождение подкорковых структур, таких как стриатума 25,26. На самом деле, специфика в формировании нейронных связей внутри и между областями мозга позволяет построение сложных систем пробирке, что улавливание многочисленные подробные проекционных систем, например, что из коры-базальных ганглиев схемы 27-30.
Через 4 - 6 недель в пробирке, одного ломтика коры и коры 31 ломтиков совместно культивировали с стриатума 26 или 32 таламус показать спонтанного вверх и вниз государств обычно встречаются в естественных условиях в уретана наркозом крысы 33. Тонкой временной организации этих государств до медведей отличительной чертой вложенных θ-и γ-колебаний свидетельствует о зрелом электрофизиологически сети пирамидных нейронов и быстро пики ГАМК-интернейронов 31. Важно отметить, что в отсутствие дофаминовых D2-рецепторов стимуляции созревания парвальбумина-положительных корковых интернейронов задерживается примерно на 2 недели в коре срез культуры 34. В соответствии с этими данными, развития учебного времени вложенных θ-, β-и γ-колебаний соответствуют, что в естественных условиях, когда кора головного мозга ломтики совместно культивировали с вентральной области покрышки (VTA), которая содержит дофаминергических нейронов проектирования до коре 6.
Эти исследования показывают, что при изучении нейронов лавины, которая решающей степени зависят от зрелых быстро ГАМК-торможения и расположены в поверхностных слоях коры 2,4, большое внимание должно быть принято для обеспечения нормального созревания коры ткани. Хотя нейронов лавины возникают в одном культур коры с течением времени курс 2 - 5 недель 4, когда требующих развития учебного времени, который согласован с развитием в естественных условиях, коры ломтиками необходимости соответствующие стимуляции дофаминовых рецепторов, например, путем совместного культивирования ломтиками коры с VTA 6.
Disclosures
Нет конфликта интересов объявлены.
Acknowledgments
Это исследование было профинансировано Отдел Внутренние программы исследований (DIRP) Национального института психического здоровья, Национального института здоровья.
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}