

Summary
一个可靠的方法来研究神经元雪崩,即规模不变的时空活动突发,在大脑皮层的临界状态动力学指示。雪崩出现自发的发展培养皮层的表面层与平面集成多电极阵列(MEA)的精确控制的条件下允许活动的长期测量。
Abstract
皮层是自发活跃,即使在没有任何特定的输入或电机输出。在发展过程中,这个活动是非常重要的皮层细胞类型,迁移和分化形成的神经元连接1。在成熟的动物,正在进行的活动,反映过去和目前的状态到感官刺激的无缝集成,计算未来行动的动物。因此,一个清醒的认识即正在进行的自发活动的组织是一个先决条件,以了解皮质功能。
众多的录音技术透露,正在进行的活动是在大脑皮层的个别活动的瞬时总结较大的事件,与细胞外微电极可以在局部领域的潜力(LFP)检测,或在脑电图(EEG),magnetoencephalogram(MEG的许多神经元组成),并从功能磁共振成像(fMRI)的大胆信号。磷酸锂铁是目前的首选方法,在学习时,在介观尺度的高时空分辨率(数千神经元)神经元群活动。在细胞外微电极,在本地同步活动空间相邻的神经元在LFP数百毫伏的快速变形的结果。当使用微电极阵列,这种变形的组织可以很方便地监控在时间和空间。
神经元雪崩描述持续2,3大脑中的神经活动的规模不变的时空组织。它们是针对4,5 体外成立6在麻醉大鼠体内的,肤浅的的皮质层,并在清醒的猴子 7 。更重要的是,理论和实证研究2,8-10表明,神经元雪崩指示的皮质精美平衡的临界状态动态,优化信息传输和信息处理。
为了研究神经元雪崩的发展,维护和调节机制, 在体外的准备工作非常有益,因为它们允许精确控制的条件下稳定的雪崩活动录音。目前的协议,介绍了如何在体外培养的神经元雪崩研究器官皮质文化,即片文化上的平面,集成的微电极阵列(MEA;见11-14)增长,利用浅层发展优势。
Protocol
1。无菌密封的玻璃商会,与MEA长期的录音
- 螺纹口玻璃瓶聚四氟乙烯塑料盖(ACE玻璃),稳固而紧密的文化室封闭所需,钢瓶被切断(Aceglass)从底部的螺纹(图1A,B)的约2毫米。水(3倍),5分钟200证明乙基酒精的沸点冲洗干净的玻璃环,让干。
- 分装芯片解决方案需要附加玻璃戒指来测量表面。混合15毫升的部分A和乙彻底Sylgard 184有机硅弹性体包,让我们坐了15分钟,以去除气泡,在1毫升分装储存在-20 ° C
- 胶玻璃环,以“千年生态系统评估”(8x8网格W /内部接地电极,电极直径30微米,200/100微米电极间的距离为鼠/鼠标)(图1A,B)。小针头注射器取1毫升的硅(23℃)。应用硅玻璃环,中心MEA玻璃环,适用于硅周围构建一个更强有力的密封环外的其他层的切割面糙米,让治愈1 - 2小时〜60℃热板。
- 在层流罩3倍去离子水,70%的酒精冲洗消毒MEA室和室盖(3 ×;最后冲洗让坐在酒精10分钟),室的10分钟的曝光和第内部紫外线灯。高压灭菌器MEA室(120 ° C,湿; 45分钟),并让干燥。
- 文化室与D -聚赖氨酸的大衣内的多边环境协定的表面。对于新的多边环境协议,而亲脂性,外套从电极网格解决方案的反复液滴的愿望。对于使用过的多边环境协议,覆盖腔底部与解决方案,吸出多余的液体,使蒸发层流罩内的无菌条件下。附加盖密封存储和将来的使用测量室。
2。器官型培养的制备及生长所需的成分
- 溶于0.9%氯化钠无菌的琼脂培养基,倒入无菌培养皿(猎鹰100 X15〜5毫米的水平),放冷,用封口膜无菌包装储存。切从固体琼脂20 × 10 × 5毫米使用块。
- 商店超级胶水,如DEVCON超级胶二,包装用70%乙醇擦拭开放前的层流罩内,以保持无菌。
- 准备加入40克的葡萄糖40毫升去离子水养殖水体(Sigma公司)50%D -葡萄糖(SIGMA超,G7528)。在2毫升分装贮存于-20 ° C
- 添加4毫升50%D -葡萄糖Gey的平衡盐溶液500毫升,阴寒(混合液体/冰晶)在使用前冷冻贿赂。
- 重组在5毫升去离子养殖水体(摇轻柔,避免形成泡沫)鸡血浆,让解决方案,解决5 - 10分钟,轻轻的漩涡,并decantate的无菌培养皿中明确的内容。无菌过滤器(0.22微米孔径过滤器;蛋白)等离子溶液,分装成cryotubes 350μL(NuncTM),储存在-20 ° C。
- 因此,无菌过滤器(0.22微米孔径过滤器),分装40μL到cryotubes(NuncTM)从牛血浆的重组凝血酶,储存于-20 ° C对于工作的解决方案,Gey W / D -葡萄糖的平衡盐溶液在375μL稀释的凝血酶溶液40μL。
- 准备混合100毫升200毫升基础培养基鹰,马血清,100毫升汉克的缓冲盐溶液4毫升50%葡萄糖2毫升200毫米L -谷氨酰胺补充400毫升培养液中。可存储4 - 8周100毫升PYREX瓶在4 ° C
- 准备通过混合0.3毫米阿糖胞苷胞嘧啶-β- D - arabinofuranoside,和0.3毫米0.3毫米尿苷,5 - 氟-2' - 脱氧尿苷,无菌过滤,分装200μL,并储存在-20 ° C 6的有丝分裂抑制剂 - 12个月。
3。皮层和腹侧被盖区(VTA)组织剥离(时间:<1小时)
- 议事产量皮层和腹侧被盖区〜12共同培养大鼠或小鼠,并准备在层流罩内,无菌条件下的组织切片。组织收集的总时间应超过1小时。
- 以健康,营养良好的仔兔(产仔数〜10;腹部的“牛奶现货”的存在),在1 - 2天(PND)产后。保持轻轻地吻一个小狗,让它挂自由,并迅速在基地的脖子,用锋利的剪刀杀头。
- 对大脑清除,清除皮肤(两个横向剪刀削减),切细眼的剪刀(1矢状中线切1冠状切开皮质/小脑交界)颅骨开放。翻转回所有4个颅骨瓣。一个削尖的锅铲,削减从正面通过嗅球,尾端下方的大脑提前锅铲。轻轻抬起的头骨和大脑让它滑入无菌,冷藏,Gey的快速冷却和临时存储解决方案。重复步骤3.2至3.3(总时间为2个多动脑:<20分钟)。
- 为了获得腹侧被盖区组织,转移到无菌,干燥的Petri使用的小铲菜的大脑。轻轻滑动每个大脑1厘米左右的横盘整理,进一步消除多余的液体。日冕,在使用刀片小脑的水平纵向切开取出脑干。
- 机械稳定在大脑切片程序的安装光盘上胶琼脂块。放置在磁盘上的琼脂块前一个几毫米(避免感人的琼脂胶),细线的强力胶。
- 使用一个小铲,转让和挂载每个大脑额极下来。确保正面两极粘在安装光盘和腹侧双方接触没有任何胶水残留的琼脂,以达到合适的机械在切割和切割片升空。稳定
- 小心淹没和大脑在vibratome托盘组件(如莱卡VT1000)的安全安装磁盘充满无菌的,冷冻Gey的解决方案。一个仔细清洗刀片(90%乙醇),降低振动频率最高的脑冠状切片和前进速度相对较低,在400〜500微米的厚度。使用一个倒置的巴斯德吸管吸球,转让和收集片含有35 × 10毫米,用无菌的,冷冻Gey的解决方案的Petri充满菜的腹侧被盖区(图1C,看到也冠状板 18 - 15 20 E22)。
- 皮质部分,重复步骤3.2 - 3.6,但适用于大脑皮层和小脑之间的纵向切开,并安装与额极forebrains。对未来的Cortex夹层约3冠状切片(厚度为350微米)的纹状体水平开始收集。
- 用微型刀的碎刀片,解剖1〜2毫米宽的额叶皮层和中脑区包含立体显微镜下腹侧被盖区(图1C)的冠状切面。收集组织切片分别在小碟充满冷冻Gey的解决方案(如玻片)。
4。安装在“千年生态系统评估”的皮层和腹侧被盖区组织切片(时间:<1小时)
- 放置于室温下一个重点的电极阵列的立体显微镜测量。血浆25μL液滴在干净,无粉尘,无菌电极阵列矩阵中央。用小铲刀,小心滑入一个皮层和腹侧被盖区部分血浆滴。
- 放在散热板测量,重新调整视图,让阴寒为〜15秒,然后添加到等离子体液滴的凝血酶25μL。使用凝血酶枪头,小心地跨越千年生态系统评估“的小圆周运动的等离子体/凝血酶的混合物。不要直接接触脆电极阵列。轻轻的位置上皮层的背侧边境沿线数组数组的第二个电极行。通过这种方式,开发浅层最终将覆盖数组的提醒。腹侧被盖区被放在相邻的腹皮层部分的边界(图1D)。
- 第松散关闭千年生态系统评估室保持高湿度,而坐在MEA /文化大会在室温〜5分钟罩内,让等离子/凝血酶凝血。同时,重复步骤4.1 - 4.3 3更多的文化。
- 小心添加600μL文化中的小水滴中的文化室,使用25 × 5 / 8针1毫升注射器。
- 紧紧贴着千年生态系统评估室和地方MEA /文化大会上的摇摆存储盘内的孵化器(图1B)。为了加快程序,3 - 4多边环境协定可在重叠的序列组装。大会时间为12多边环境协定应<1小时。
- 在体外 (DIV),后2天,加10μL的有丝分裂抑制剂。文化传媒刷新了60%,在4格,以后每4天。
5。电生理记录和激励生成
- 在局部领域的潜力(LFP)和消防动作电位神经元的倾向显着变形之间的关系,建立后〜1 个星期5,6纪录在24 kHz〜10分钟的自发活动,从每个电极的MEA(五金:MEA1060 W /消隐电路,X1200增益,12位A / D,0-4096 mV的范围内,多声道系统;软件:MC_Rack)。理由是通过内部的接地电极,或外部加入的Ag / AgCl的半细胞。
- 单独用一个带通滤波器1 LFP - 200赫兹(带通300 - 3000赫兹)从外秒杀活动。秒杀活动可以进一步分为单级和多单元使用脱线扣球的分拣机(如Plexon公司公司)的活动。计算油价平均为每个电极。皮质文化,最平均,在培养的神经元扣球的首选时间确定为负LFP挠度(nLFP)。
- 从大幅面痕迹噪声-3标准偏差(SD)的阈值计算每个电极(图2)确定nLFPs的高峰时间和振幅,跨门槛(图2B,C)。选择一个时间斌ΔT(例如介于2 - 8毫秒),并识别连接所有电极在连续的时间长度为ΔTbines(图2D相同nLFPs的阵列上的时空nLFP集群;有关详细信息 ,看到2,4 ,5)。
- 为了确定神经元雪崩,计算每个nLFP集群的大小,活跃电极或nLFP幅度的总和,例如,建造一个大小直方图,在双对数坐标图。对于神经元雪崩,粒度分布如下功法在双对数坐标2(图2E,F)的直线近似。法律对权力的统计测试,请参见16。
- 引出选择电极,通过它的电流控制幅度小号刺激应用(STG 1008刺激发生器,多声道系统),在组织中引起的反应。为了减少电极损坏,使用范围有限,电中性刺激与双极方波单冲击:采用振幅S 50微秒100微秒幅度+ S / 2和S之间的10 - 200微安。见主人的进一步细节手册。
- 要记录的动态范围记录LFP的反应,在4 kHz的采样率在所有电极刺激后500毫秒,记录刺激。消隐电路(多通道系统)断开时的刺激,减少刺激的文物和防止前级放大器饱和头级放大器使用。
6。代表性的成果:
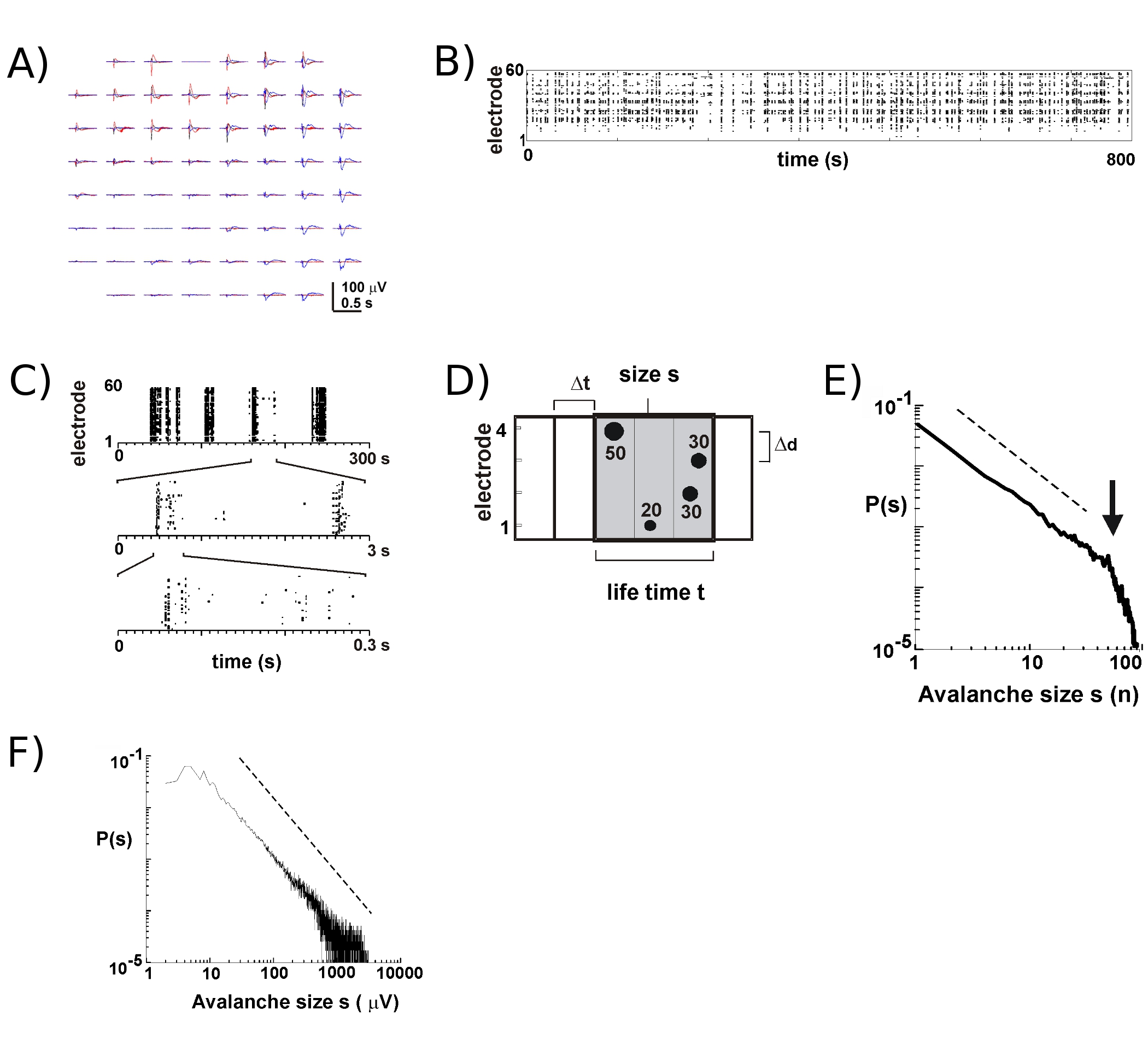
随着新的多边环境协定的约8 - 9 10文化生存的许多个星期。我们长期的录音大多培养箱内培养液中,这使我们能够遵循个别文化的发展,在许多个星期的课程5。根据我们的实验中,大幅面录音可以可靠地获得超过100天文化的多边环境协定。相比之下,外秒杀活动是更可靠的测量与相对新的多边环境协定(<40天文化)。在一个典型的实验中,我们转移到存储盘(图1B,右)多边环境协定的托盘头阶段附加保持密封的文化室(图1B,左)。皮层5,皮质的腹侧被盖区共同文化6,以及在麻醉大鼠和清醒的猴子体内 7,在表面层的神经元放电神经元雪崩期间主要发生密切的磷酸锂铁的峰值负偏转( nLFP )。因此,本地同步的神经元群体的时空组织,可以通过测量发生在空间和时间上的阵列17 nLFPs的估计。
“千年生态系统评估”上的活动,往往出现在时间上的集群,这样,在一个电极的活动是伴有其他部位的活动。在这样的活动期间LFP典型波形如图2a所示,超过绘制3集群,除了发生几秒钟。对于每一个群集,负磁场偏转可以看出几个电极内窗口1号当提取nLFP高峰跨多个负面的SD阈值,nLFP高峰时段的形式的活动,是在“列”的点附近重合nLFPs各种焊条(图2B)的代表在其中一个光栅方便的可视化。这一活动的时空组织是相当复杂的,“列”,从而出现更多或更少的时间分辨率低的同质,组成独立的集群,在更高的时间分辨率等(图2C)。事实上,时空nLFP集群的出现,是高度组织化的皮质网络。更具体地说,该组织是神经元雪崩的规模不变。这表现在给定的时间分辨率ΔT计算的簇大小的概率。在这里,集群组成,在相同或连续的时间箱(图2D)发生的nLFPs。当这种集群的大小总数nLFPs,每个集群或综合nLFP每个集群振幅表示,簇的大小分布,揭示了一个功法,其斜坡已被证明是 -1.5 2,4,5,7(图2E,F)。请注意,这种分配标识的簇大小,规格有S kxs,其中k是一个常数因子的比例尺度不变的订购, 是 k -1.5,这是独立的S.此功法组织是独立的阵列2的大小,ΔT2,时间分辨率和阈值,以找出重要nLFP的偏差 。由于与神经元组的大小7 nLFP幅度尺度,nLFPs规模不变的组织反映了规模不变,即分形,订货Øf本地同步,包括各种规模的神经元组。

图1(A)“千年生态系统评估”的侧面和顶部的意见安装螺纹玻璃环,和相应的上限。 (二)孵化器的内部。左:探头安装孵化器的条件下,允许从一个单一的文化录制。右:托盘,文化的发展的许多多边环境协定。侧车轮:步进电机控制设备摇摆交替淹没,暴露于大气的阶段,文化的发展所需。 (三)用于皮质的腹侧被盖区共培养大鼠冠状片示意图。皮质部分(左)和中脑的部分(中,右)中含有的腹侧被盖腹侧被盖区(VTA;灰色),获得沿虚线切割。 CTX:皮质; WM:白色物质; CPU:纹状体; VTA:脑桥:脑桥区。也相应冠状板8,18和20 15。 (四)安置和一个单一的皮层腹侧被盖区上的多边环境协定和其对第9格文化发展联合培养增长。注意的扁平化的文化和阵列上逐步扩大。反光组织部分表明退化的细胞和组织碎片。健康的组织,是不透明的,可见光透射下的灰。

图(2)神经元在大脑皮层器官文化的雪崩。 (一)三个阵列上的自发活动期间Overplot,几秒钟分开。请注意,每个活动期间,许多电极阵列上的负面LFP挠度(每种颜色的标签,一个活动期)组成。 (二)每个电极nLFPs的负峰值时间是组装成一个活动的栅格。 “表示Column'状结构附近的同步活动的时期。 (三)需要注意的是出现在一个时间尺度高度同步的列较高的时间尺度(时间尺度3)的多个列组成。 (四)神经元雪崩算法的示意图。在一个2 × 2电极阵列高峰时间和幅度的负LFP挠度过路- X SD的噪声阈值(nLFP)确定。 nLFPs时空组织聚成先后活跃的时间宽度ΔT箱。簇大小是确定的活性位点,即与nLFP电极(S = 4),或综合nLFP振幅总和(S = 130μV)。续航时间是衡量ΔT的倍数。 (E,F)簇大小分布的功法确定为神经元雪崩的集群。请注意,δD为阵列(这里为200微米)的一个特定的极间距离的选择,引入了一个特别的ΔT的动态时,应遵守。更具体地说,在其中的比例ΔD/ΔT接近平均在网络中的传播速度,在这功法的坡度α神经元雪崩 2,4,5接近-1.5 。请点击这里看大图图2。
Discussion
1。技术问题:
- 无菌技术。多边环境协定和文化准备的准备工作都是层流罩内,在无菌条件下进行。不使用抗生素,从而影响神经元的活动,在任何时间,在编制和文化的过程中。
- 等离子/凝血酶凝血和组织上坚持对MEA 多边环境协定。组织的生存需要一个谨慎的平衡之间的等离子/凝血酶凝血和组织暴露在大气中时所需的时间。一个凝血时间短,风险过早支队从MEA部分,而长时间暴露到大气中的触发器组织变性。由于凝血酶解决方案的实力,决定了凝血过程的速度,这是一个非常重要的参数,成功,健康的文化MEA表面附着。我们取得最佳效果,使用1000个单位(1千片; 1美国国立卫生研究院单位= 0.324 ± 0.073毫克)。更重要的是,不完整的空间异质性凝血血浆/凝血酶的解决方案,促进在培养等离子体破裂的结果混合。这些等离子的“孔”文化健康造成严重影响,从微电极分离的文化,从而损害的电生理记录的质量。冷却板,在多边环境协定/组织装配工作,减缓凝血,并允许为均质混合血浆/凝血酶的解决方案和正确定位组织切片。
同样,加入单液滴中文化室淹没后仅5分钟凝固的文化,大大减少了由于表面张力的组织分队的风险。一个成功的文化,将扁平化“千年生态系统评估,并略有扩大显示周的健康成长,没有任何不完整的组织,多边环境协定的联系或组织变性(例如图1D)主要标志。 - 组织剥离。微型刀,极大地提高了我们的清扫过程。我们使用双重优势刀片(精细的科学工具 - 易碎的手术刀片 - 100050-00),从中我们分裂〜2毫米宽的“刀片”用钳子,和他们用手术刀持有人持有。从冠状切片用刀片垂直平稳,公司的议案解剖的组织样本,从而大大降低了由于机械应力拉动整个组织,提高文化健康。
- 组织冷却。适当的散热片和组织切片在准备文化的成功至关重要。我们使用佩尔蒂埃元素连接到一个金属圆盘下方内置的定制冷板。佩尔蒂埃元素产生的热量是通过冷水灌注中删除。这大大减少了准备时间和标准化,在每个阶段的准备冷却(DOLD实验室与工程博士131种植塞甘,TX 78155,(830)560-1471))。
- 孵化条件。一个定制孵化器与精确的内部摇摆条件下,我们在培养上的多边环境协定的片的成功至关重要。内部的摇摆设备的基础上由Multichannelsystems原有的内部设计(目前市面上的),包括连接到两个大的一侧车轮的托盘。步进电机和电脑控制允许为一个精确的摇摆轨迹(摇摆角,摇摆速度,间歇性停顿)。最终,片文化需要公开的氛围和文化,慢交替的媒介。传统的方法是放置到狭窄的管片,慢慢地沿着有最长的轴旋转文化。在这里,自转速度慢不产生机械应力,由于旋转本身,和旋转速度足够高,以获得最佳的“进料/呼吸周期约5 - 10分钟的持续时间。千年生态系统评估室,其总量的〜2毫升,小的培养基量为中小型空调需要由组织本身更紧凑的内部,构成一个巨大的挑战。 ±〜70 °角(周期时间:200秒)之间的摇摆“千年生态系统评估”的液相和大气之间的文化转换的摇摆速度减慢,并停止在极端的角度,长时间暴露在大气中的摇摆已对于文化的生存至关重要。
2。皮质文化的发育年龄来研究神经元雪崩
通常采取的PND 0 - 1片从大鼠皮层,急性和培养“千年生态系统评估”的几个星期。早期的研究已清楚地表明,单皮质切片培养, 在体外培养的几个星期后,保持与识别细胞类型的层状结构,可以很容易地比较在体内细胞类18,18-21。分层组织已在体外系统在此方便地用于研究丘脑22日至24日在开发过程中,以及支配的皮层皮层下结构,如纹状体 25,26驾驶。事实上,在形成的大脑区域内和跨的神经元连接的特殊性, 允许复杂的体外系统,重新夺回了许多详细的投影系统的建设 ,如皮层基底节电路27-30。
经过4 - 6周在体外,单皮质片31和皮质切片培养与纹状体26或32丘脑显示自发向上和向下通常在33乌拉坦麻醉大鼠体内发现的状态这些最多的国家的优秀时间组织负有标志嵌套θ-和γ-振荡的锥体神经元和GABA能interneurons快速扣球31 electrophysiologically成熟的网络指标。重要的是,在多巴胺D2受体的刺激的情况下,成熟的小白蛋白阳性皮质interneurons延迟约2 周皮质切片文化34。嵌套在这些研究结果,发展的时间当然θ,β-和γ-振荡匹配体内的皮质切片时,腹侧被盖区(VTA),其中包含的多巴胺能神经元投射到联合培养皮质6。
这些研究表明,在研究神经雪崩,关键取决于成熟快的GABA能抑制,并在位于大脑皮层2,4的 浅层时,应采取非常谨慎,以保证适当的皮层组织成熟。虽然神经元雪崩出现超过2的时间当然在单一皮质文化- 5 周 4,当需要一个发展的时间当然这是在体内发育匹配,皮质片需要适当的多巴胺受体刺激,例如共培养皮层切片腹侧被盖区6。
Disclosures
没有利益冲突的声明。
Acknowledgments
这项研究是由院内研究计划部的国立精神卫生研究所,国立卫生研究院(DIRP)。
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}