

Summary
Una forma sólida para estudiar las avalanchas neuronales, es decir, invariante en escala espacio-temporal de ráfagas de actividad, lo que indica la dinámica de estado crítico en la corteza. Avalanchas surgen espontáneamente en el desarrollo de las capas superficiales de la corteza cultivo que permite mediciones a largo plazo de la actividad con planos las redes distribuidas multi-electrodo (MEA) en condiciones controladas con precisión.
Abstract
La corteza es espontáneamente activo, incluso en ausencia de cualquier entrada o salida del motor. Durante el desarrollo, esta actividad es importante para la migración y diferenciación de los tipos de células corteza y la formación de conexiones neuronales 1. En el animal adulto, la actividad en curso refleja el pasado y el actual estado de un animal en la que los estímulos sensoriales están perfectamente integrados para calcular las futuras acciones. Por lo tanto, una clara comprensión de la organización del curso la actividad espontánea es decir, es un requisito previo para comprender la función de la corteza.
Numerosas técnicas de grabación reveló que la actividad en curso en la corteza se compone de muchas neuronas, cuyas actividades individuales transitoriamente suma a los grandes eventos que se pueden detectar en el potencial del campo local (LFP) con microelectrodos extracelulares, o en el electroencefalograma (EEG), la magnetoencefalograma (MEG ), y la señal BOLD de resonancia magnética funcional (fMRI). La LFP es actualmente el método de elección para estudiar la actividad neuronal de la población con una alta resolución temporal y espacial en la escala mesoscópica (varios miles de neuronas). En el microelectrodo extracelular, a nivel local sincronizado actividades de resultado espacialmente neighbored neuronas en desviaciones rápido en la LFP hasta varios cientos de microvoltios. Cuando se utiliza una serie de microelectrodos, las organizaciones de tales desviaciones pueden ser convenientemente controlados en el espacio y el tiempo.
Avalanchas neuronales describir la organización espacio-temporal invariante en escala de la actividad en curso neuronal en el cerebro 2,3. Que son específicos de las capas superficiales de la corteza a lo establecido en 4,5 in vitro, in vivo en ratas anestesiadas 6, y en el mono despierto 7. Es importante destacar que los estudios teóricos y empíricos sugieren que 2,8-10 avalanchas neuronales indican una dinámica crítica exquisitamente equilibrado estado de la corteza que optimiza la transferencia de información y procesamiento de la información.
Con el fin de estudiar los mecanismos de desarrollo avalancha neuronal, el mantenimiento y la regulación, en preparaciones in vitro son muy beneficiosos, ya que permiten grabaciones de actividad estable avalancha en condiciones controladas con precisión. El actual protocolo describe la forma de estudiar las avalanchas neuronales in vitro mediante el aprovechamiento de desarrollo de la capa superficial de la corteza organotípicos culturas, las culturas, es decir corte, crecido en matrices de plano, de microelectrodos integrados (MEA, ver también 11-14).
Protocol
1. Cámara estéril, de vidrio con cierre de MEA registros a largo plazo
- Cilindros de vidrio con rosca de teflón de plástico tapa (Ace Glass), necesarias para la clausura Cámara de Cultura del seguro y firme, se cortan (Aceglass) de aproximadamente 2 mm de fondo de la rosca (Fig. 1A, B). Anillos de limpiar el cristal de un enjuague con agua (3x) y hervir durante 5 minutos en la prueba 200 etil-alcohol, dejar secar.
- Solución alícuota de silicio para conectar anillos de vidrio a la superficie de los AMUMA. Mezcla de 15 ml de las partes A y B de Sylgard 184 Kit de elastómero de silicona completamente, deje reposar durante 15 minutos para eliminar las burbujas de aire, almacenar en alícuotas de 1 ml a -20 ° C.
- Cola de anillo de cristal en el AMUMA (8x8 red w / electrodo interno, 30 m diámetro del electrodo, 200/100 m distancia entre los electrodos de rata / ratón) (Fig. 1A, B). Tomar 1 ml de silicio (23 ° C) en la jeringa con aguja de pequeño calibre. Aplique silicona para pulir la superficie de corte del anillo de cristal, el anillo central de vidrio de MEA, aplique una capa adicional de silicona en la parte exterior del anillo para un sello más fuerte, que la curación por 1 - 2 horas a 60 º C en un plato caliente.
- MEA esterilizar la cámara y las tapas de la cámara en una campana de flujo laminar de lavado 3 veces en agua desionizada seguido por alcohol al 70% (3 x, por último aclarado deje reposar durante 10 minutos en alcohol), seguido por la exposición de 10 minutos de la cámara y el interior de la PAC a la luz UV . Autoclave MEA cámara (120 ° C, húmedo, min 45) y dejar secar.
- Escudo MEA superficie interior de la cámara de cultivo con poli-D-lisina. De los acuerdos ambientales multilaterales nuevos, que son bastante lipofílica, capa por aspiración repetida gota de la solución de la red de electrodos. Para utilizar los acuerdos ambientales multilaterales, cubierta con una solución de fondo de la cámara, aspirar el exceso de líquido, permita que se evapore en condiciones de esterilidad dentro de campana laminar. Adjuntar tapa para sellar la cámara de MEA para su almacenamiento y su uso futuro.
2. Ingredientes necesarios para la preparación y el crecimiento de cultivos organotípicos
- Disolver el agar estéril de NaCl al 0,9%, se vierte en placa de Petri estéril (Falcon, 100 x15; ~ 5 mm de nivel), dejar enfriar y envolver estéril con Parafilm para su almacenamiento. Reducción del 20 x 10 x 5 mm bloques de agar sólido para su uso.
- Tienda de pegamento, por ejemplo, Devcon Super Glue II, con el embalaje limpiados con EtOH al 70% antes de la apertura, dentro de la campana de flujo laminar para mantener la esterilidad.
- Prepare un 50% de D-glucosa (SIGMA ultra, G7528), añadiendo 40 g de glucosa a 40 ml de agua desionizada cultura (Sigma). Almacenar en alícuotas de 2 ml a -20 ° C.
- Agregar 4 ml de 50% de D-glucosa a 500 ml de solución salina equilibrada de Gey, y la frialdad de lodo (mezcla de cristales líquidos / hielo) en el congelador antes de su uso.
- Reconstituir plasma de pollo en 5 ml de agua de cultivo de-ionizada (agitar suave, evitar la formación de burbujas), permita que la solución reposar durante 5 - 10 min, agitar suavemente, y decántate el contenido claro en una placa de Petri estéril. Estériles con filtro (filtro de 0,22 micras de poro; proteína) solución de plasma, alícuota de 350 l en criotubos (NuncTM), almacenar a -20 ° C.
- Reconstituir la trombina a partir de plasma de bovino en consecuencia, estéril-filtro (filtro de 0,22 micras de poro), alícuota de 40 ml en criotubos (NuncTM), almacenar a -20 ° C. Para la solución de trabajo, diluir 40 ml de la solución de trombina en 375 l de solución salina equilibrada de Gey w / D-glucosa.
- Preparar 400 ml de medio de cultivo mediante la mezcla de 100 ml de suero de caballo, 200 ml basal medio de Eagle, 100 ml de Hank solución salina a la que se añaden 4 ml de glucosa al 50% y 2 ml de 200 mM L-glutamina. Se pueden almacenar 4 - 8 semanas en botellas de 100 ml PYREX a 4 ° C.
- Prepare inhibidor de la mitosis mediante la mezcla de 0.3 mM uridina, 0,3 mM mM ARA-C de citosina-β-D-arabinofuranoside, y 0,3 5-fluoro-2'-desoxiuridina, filtro estéril, l alícuota 200 y almacenar a -20 ° C durante 6 - 12 meses.
3. La corteza y el área tegmental ventral (VTA) la disección de tejidos (tiempo: <1 hora)
- Los rendimientos de Procedimiento corteza y las secciones de tejido para VTA ~ 12 co-cultivos de ratas o ratones, y se prepara dentro de una campana de flujo laminar bajo condiciones estériles. El tiempo total para la recogida de tejidos deben ser de 1 hora.
- Tener cachorros sanos, bien nutridos (tamaño de la camada ~ 10, la presencia de un "punto de leche abdominal) en 1 - 2 días después del parto (PND). Mantenga un cachorro suavemente por el hocico, permita que cuelgue libremente y con rapidez decapitar en la base del cuello con unas tijeras afiladas.
- Para la extracción del cerebro, quitar la piel (dos cortes de tijera lateral), cortar con tijeras cráneo abierto los ojos bien (1 corte medio sagital, coronal un corte en la corteza / de conexiones cerebelo). Voltear hacia atrás las 4 aletas cráneo. Con una espátula afilada, corte frontal a través del bulbo olfatorio, una espátula avanzar caudalmente por debajo del cerebro. Con cuidado, levante el cerebro fuera del cráneo y se deja caer en estériles, refrigeradas, Gey solución para un enfriamiento rápido y el almacenamiento temporal. Repita los pasos del 3,2 al 3,3 por 2 cerebros más (tiempo total: <20 min).
- Para obtener VTAtejido, la transferencia de los cerebros en una fuente estéril, seco Petri usando una pequeña espátula. Suprimir el exceso de líquido deslizando suavemente cada cerebro alrededor de 1 cm de lado. Retire del tallo cerebral por un corte coronal, vertical a nivel del cerebelo con una cuchilla de afeitar.
- Pegamento de bloques de agar para el montaje de disco para la estabilización mecánica del cerebro durante el corte de procedimiento. Coloque una línea fina de pegamento de unos pocos milímetros por delante del bloque de agar en el disco (evitar el contacto con la goma de agar).
- Usando una espátula pequeña, la transferencia y montaje de cada cerebro, polo frontal hacia abajo. Asegúrese de que los polos frontales están pegados a discos de montaje y que los lados ventral tocar el agar sin residuos de pegamento con el fin de lograr la estabilización de las propiedades mecánicas durante el corte y fácil de elevación inicial de las rebanadas cortadas.
- Con cuidado, sumergirse y disco de montaje seguro con el montaje del cerebro en una cubeta vibratome (por ejemplo, Leica VT1000) llena de estéril, refrigerados Gey solución. Con una hoja de afeitar limpia cuidadosamente (90% EtOH), corte cortes coronales del cerebro medio a la frecuencia de vibración más alta y la velocidad de avance relativamente baja con un espesor de 400 500 micras. Usando una pipeta Pasteur invertida con pera de succión, la transferencia y recoger los trozos que contiene el VTA de 35 x 10 mm placas de Petri llenas de estériles, refrigeradas Gey solución (Fig. 1C, ver también la placa de la corona 18 a 20 en E22 en 15).
- De la corteza secciones, repita los pasos 3,2 a 3,6, pero se aplican corte vertical entre la corteza y el cerebelo, y montar prosencéfalos con polo frontal hacia arriba. Alrededor de 3 cortes coronales (350 micras de espesor) a partir del nivel del cuerpo estriado se recogen para la disección de la corteza en el futuro.
- Con un cuchillo de micro-hechas de hojas de afeitar rota, diseccionar ~ 2 mm de ancho sección coronal de la corteza frontal y áreas del cerebro medio que contiene el VTA (Fig. 1C) bajo un microscopio estereoscópico. Recoger los cortes de tejido por separado en pequeños platos (por ejemplo, la cámara de diapositivas) lleno de solución fría de Gey.
4. Montaje de la corteza y VTA Rebanadas de tejido en el MEA (Hora: <1 hora)
- Posición MEA a temperatura ambiente bajo un microscopio estereoscópico con la matriz de electrodos en el foco. Centro de una gota de 25 l de plasma en la limpia, libre de polvo, y la matriz estéril electrodo matriz. Usando espátulas pequeñas, deslice con cuidado la corteza y la sección de VTA en la gota de plasma.
- MEA lugar en la placa de enfriamiento, reorientar punto de vista, dejar que enfríe por unos 15 s, a continuación, añadir 25 l de trombina en la gota de plasma. Con la punta de pipeta trombina, extiende con cuidado la mezcla de plasma / trombina con pequeños movimientos circulares a través de la MEA. No toque la guía de electrodos directamente frágil. Suavemente la posición de la corteza de la matriz con el borde dorsal a lo largo de la fila segundo electrodo de la matriz. De esta manera, las capas superficiales de desarrollo eventualmente cubrir el recordatorio de la matriz. El VTA se coloca al lado del borde ventral de la sección de la corteza (Fig. 1D).
- Cap y suelta cerca de la cámara de MEA para retener la humedad alta, mientras que el conjunto de MEA / cultura se encuentra en ~ 5 minutos dentro de la campana a temperatura ambiente para permitir el plasma / trombina coagulación. Mientras tanto, repita los pasos 4,1 a 4,3 por 3 culturas más.
- Cuidadosamente añadir 600 l de medio de cultivo en pequeñas gotas para la cámara de cultivo con una jeringa de 1 cc con 25 x 5 / 8 de aguja.
- Cierre bien la cámara de MEA y MEA lugar / montaje cultura en la bandeja de almacenamiento de rock dentro de la incubadora (Fig. 1B). Para acelerar el procedimiento, 3-4 AMUMA pueden ser ensamblados en secuencias que se solapan. Tiempo de montaje de 12 acuerdos ambientales multilaterales debería ser <1 hora.
- Después de 2 días in vitro (DIV), añadir 10 l de inhibidor de la mitosis. Actualizar los medios de cultivo en un 60% a los 4 DIV y cada 4 días después.
5. Registro electrofisiológico y generación de estímulos
- Para establecer la relación entre las desviaciones significativas en el potencial del campo local (LFP) y la tendencia de las neuronas disparen los potenciales de acción, después de aproximadamente una semana la actividad de registro de 5,6 espontánea a los 24 kHz de ~ 10 minutos de cada electrodo de la MEA (Hardware : MEA1060 w / circuito de cierre, x1200 ganancia de 12 bits A / D, rango 0-4096 mV, sistemas multicanal; Software: MC_Rack). Tierra se proporciona a través del electrodo de masa interna o externamente mediante la adición de una célula de Ag / AgCl y medio.
- Separar la LFP con un filtro de paso de banda de 1 a 200 Hz de la actividad extracelular pico (de paso de banda 300 - 3000 Hz). Actividad pico pueden ser clasificados en la actividad individual y de varias unidades fuera de línea utilizando clasificadores de pico (por ejemplo, Plexon Inc.). Calcular repunte provocado promedios para cada electrodo. Para las culturas corteza, la mayoría de los promedios identificar desviaciones negativas LFP (nLFP) como la hora preferida de picos neuronal en la cultura.
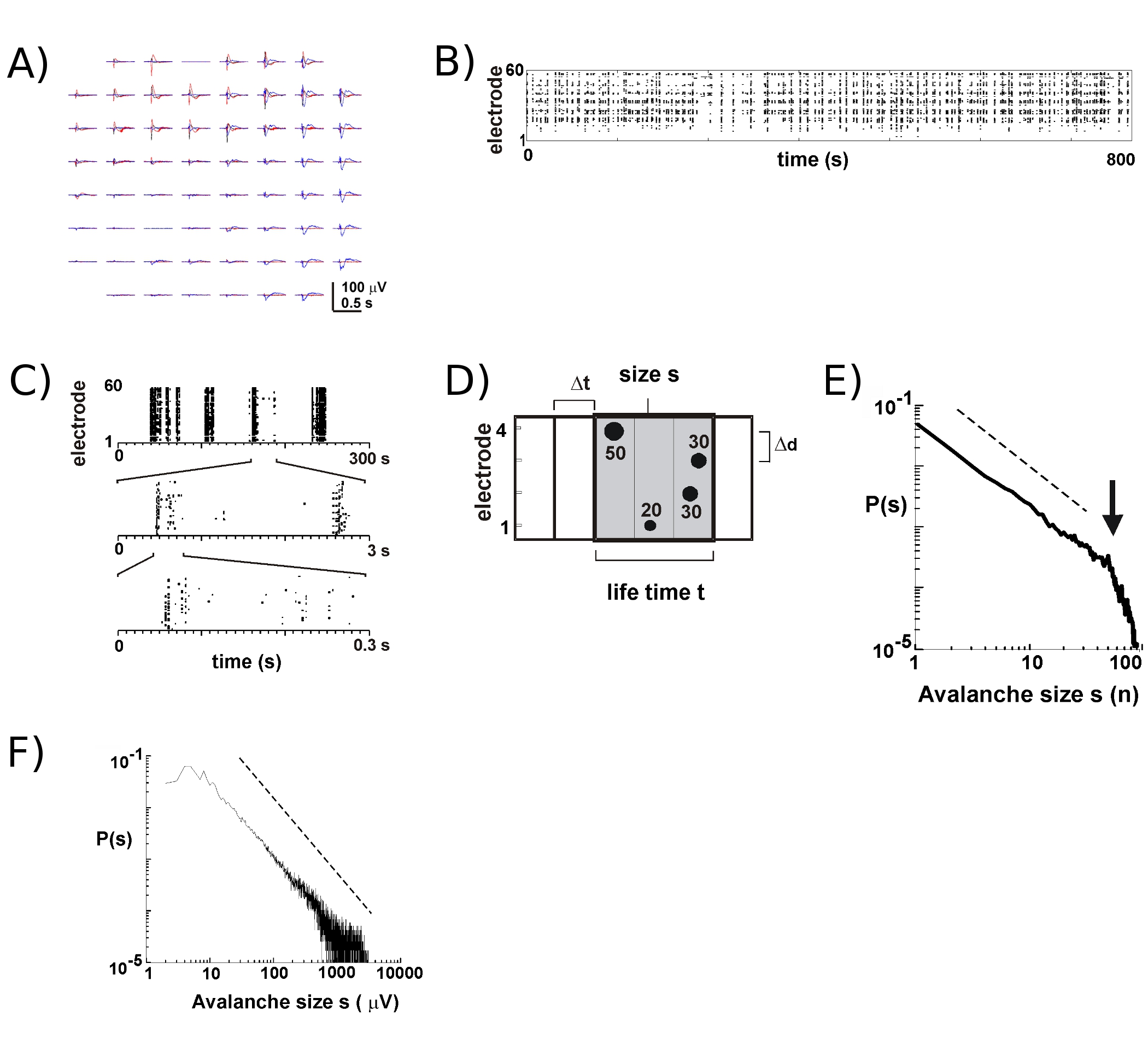
- Se calcula para cada electrodo un umbral de -3 desviaciones estándar del ruido (SD) de las huellas LFP (Fig. 2), Determinar las horas punta y amplitudes de nLFPs que cruzan el umbral (Fig. 2B, C). Elija un recipiente tiempo Dt (por ejemplo, entre 2 a 8 ms) e identificar grupos espaciotemporales nLFP en la matriz mediante la concatenación de todos los nLFPs electrodos que están en el mismo tiempo combina sucesivas Dt longitud (Fig. 2D, para más detalles véase 2,4 , 5).
- Con el fin de identificar las avalanchas neuronales, calcular el tamaño de cada grupo nLFP, por ejemplo, número de electrodos activos o la suma de las amplitudes nLFP, construir un histograma de tamaño, y la trama de doble logarítmica coordenadas. Las avalanchas neuronales, la distribución del tamaño sigue una ley de potencia aproximada por una línea recta en doble logarítmica dos coordenadas (Fig. 2E, F). Ver 16 para las pruebas estadísticas sobre las leyes de alimentación.
- Obtener las respuestas evocadas en el tejido mediante la selección de un electrodo a través del cual los estímulos de corriente regulada con amplitud S se aplican (generador de estímulos STG 1008, los sistemas multicanal). Para reducir el daño de los electrodos, el uso de un rango limitado, de carga neutra la estimulación de los choques individuales con onda cuadrada bipolar: 50 ms con amplitud-S, seguido de 100 ms con una amplitud de + S / 2 y S entre 10 a 200 mA. Consulte el manual del propietario para más detalles.
- Para grabar el rango dinámico 9 de estímulo récord registraron las respuestas de LFP en 4 kHz frecuencia de muestreo en todos los electrodos después de 500 ms después de la estimulación. El uso de circuitos de cierre (sistemas multi-canal) que desconecta los amplificadores de la etapa en la cabeza durante la estimulación para reducir los artefactos de estímulo y para prevenir pre-amplificador de saturación.
6. Los resultados representativos:
Con los acuerdos ambientales multilaterales nuevos de 8 - 9 de cada 10 culturas sobrevivirán por muchas semanas. La mayoría de nuestros registros a largo plazo llevará a cabo dentro de la incubadora en medio de cultivo, lo que nos permite seguir el desarrollo de las culturas individuales a lo largo de varias semanas 5. Con base en nuestros experimentos, grabaciones LFP puede ser fiable obtenidos con los acuerdos ambientales multilaterales utilizado por más de 100 días de cultivo. Por el contrario, la actividad extracelular pico es más fiable con los acuerdos ambientales multilaterales relativamente nueva (<40 días de cultivo). En un experimento típico, que la transferencia de un acuerdo multilateral de la bandeja de almacenamiento (Fig. 1B, derecha) para la bandeja con la cabeza unida etapa (Fig. 1B, a la izquierda) el mantenimiento de la cámara de cultivo cerrado. De la corteza 5, la corteza-VTA co-cultivos 6, así como en la rata anestesiada 6 y el mono despierto en vivo 7, la descarga neuronal durante las avalanchas neuronal en las capas superficiales se produce principalmente cerca de la desviación de pico negativo de la LFP (nLFP ). Así, la organización espacio-temporal de local sincronizado grupos neuronales se puede estimar mediante la medición de la ocurrencia de nLFPs en el espacio y el tiempo en los 17 matriz.
La actividad en la MEA tiende a surgir en agrupamientos temporales, de tal manera que la actividad en uno de los electrodos se acompaña de la actividad en otros sitios. Formas de onda típica de la LFP durante los períodos de actividad se muestran en la figura 2A, por más de tres grupos de trazado se producen unos segundos de diferencia. Para cada grupo, las desviaciones negativas de campo se puede ver en varios electrodos dentro de una ventana de 1 s. Al extraer los picos nLFP que cruzar un umbral de múltiples negativos SD, la actividad en la forma de nLFP las horas pico está muy bien visualizado en una trama en la que "columnas" de puntos representan cerca nLFPs coincidente en varios electrodos (Fig. 2B). La organización espacio-temporal de esta actividad es bastante compleja, "columnas", que aparecen más o menos homogénea en baja resolución temporal, se componen de distintas agrupaciones con mayor resolución temporal y así sucesivamente (Fig. 2C). De hecho, el surgimiento de grupos espaciotemporales nLFP está muy bien organizada en redes corticales. Más específicamente, la organización es invariante en escala de avalanchas neuronales. Esto se demuestra mediante el cálculo de la probabilidad de tamaños de clúster en un Dt resolución dada temporal. Aquí, los grupos se componen de nLFPs que se producen en los contenedores vez o sucesivas (Fig. 2D). Cuando el tamaño de este grupo se expresa en el número total de nLFPs por racimo, o integrado amplitudes nLFP por racimo, las distribuciones de tamaño de clúster revela una ley de potencia, cuya pendiente se ha demostrado que se -1,5 2,4,5,7 ( fig. 2E, F). Tenga en cuenta que esta distribución se identifica un orden invariante en escala de tamaños de clúster que es la relación entre los tamaños de KXS s, donde k es una constante, es de -1.5 k, que es independiente de s. Esta organización de ley de potencia es independiente del tamaño de la matriz 2, la resolución temporal Dt 2, y el umbral para identificar desviaciones significativas nLFP 7. Debido a la amplitud de las escalas nLFP con el tamaño del grupo neuronal 7, la organización invariante en escala de nLFPs refleja una invariante en escala, es decir, fractal, ordenar of localmente sincronizados grupos neuronales que incluyen todos los tamaños.

Figura 1. (A) Vistas laterales y parte superior de la AMA con el anillo de vidrio roscado montado, y la tapa correspondiente. (B) Vista interior de la incubadora. Izquierda: headstage montaje permite la grabación de una sola cultura en condiciones incubadora. Derecha: bandeja con los acuerdos ambientales multilaterales para el crecimiento de numerosos cultura. Ruedas laterales: motor paso a paso controlado por el dispositivo oscilante para alternar la fase sumergida y la atmósfera expuestos requeridos para el crecimiento de la cultura. (C) Esquema de las rebanadas de la corona utilizada para la corteza-VTA co-cultivos. Secciones de la corteza (izquierda) y secciones del cerebro medio (centro, derecha) que contiene el VTA área tegmental ventral (VTA, gris) se obtienen mediante la reducción a lo largo de las líneas de trazos. CTX: la corteza; wm: la sustancia blanca; CPU: cuerpo estriado; vta: Pons: zona del puente. Ver también las placas correspondientes coronal 8, 18, y 20 por 15. (D) Colocación y crecimiento de una sola corteza-VTA co-cultivos en el MEA y su desarrollo durante los primeros nueve DIV en la cultura. Tenga en cuenta el achatamiento de la cultura y la expansión progresiva de la matriz. Partes del tejido reflectante indican células degeneradas y restos de tejido. El tejido sano es opaco y grisáceo con transiluminación con luz visible.

Figura 2. Avalanchas neuronales corticales en cultivos organotípicos. (A) Overplot de tres períodos de actividad espontánea de la matriz, separados por varios segundos. Tenga en cuenta que cada período de actividad consiste en la LFP desviaciones negativas en muchos electrodos en la matriz (cada color etiquetas de un período de actividad). (B) veces negativos pico de nLFPs de cada electrodo se ensamblan en una trama de la actividad. "Como Column' las estructuras indicadas períodos de actividad sincrónica cerca. (C) Tenga en cuenta que las columnas que aparecen altamente sincronizada a escala de una sola vez constará de varias columnas en las escalas superior temporal (3 escalas temporales se muestra). (D) Representación esquemática del algoritmo avalancha neuronal. En un 2 x 2 el tiempo máximo de electrodos de matriz y la amplitud de las desviaciones negativas LFP (nLFP) cruzar el umbral de x SD del ruido se identifican. Organización espacio-temporal de nLFPs se agrupan en contenedores de tiempo Dt sucesivamente activa de ancho. El tamaño de un grupo es identificado por el número de sitios activos, es decir, con electrodos nLFP (s = 4), o la suma integrada de las amplitudes nLFP (s = 130 mV). El tiempo de vida se mide en múltiplos de Dt. (E, F) la ley de energía en la distribución de tamaño de cluster identifica grupos como avalanchas neuronales. Tenga en cuenta que la elección de la distancia entre los electrodos en particular Δd para la matriz (en este caso 200 micras), introduce un Dt particular en el cual la dinámica debe ser observada. Más específicamente, la proporción en que Δd / Dt se aproxima a la velocidad de propagación promedio en la red, en la que la α pendiente de la ley de potencia se aproxima a -1,5 por avalanchas neuronales 2,4,5. Por favor, haga clic aquí para ver una versión ampliada de la figura 2.
Discussion
1. Cuestiones técnicas:
- Una técnica estéril. La preparación de los acuerdos ambientales multilaterales y la preparación de la cultura son todas realizadas dentro de una campana de flujo laminar bajo condiciones estériles. Los antibióticos, que afectan a la actividad neuronal, no se utilizan en cualquier momento durante el proceso de preparación y la cultura.
- Plasma / trombina y la coagulación de la adhesión del tejido en la supervivencia de los AMUMA. Tejido en el MEA requiere un cuidadoso equilibrio entre el tiempo requerido para el plasma / trombina coagulación y el tiempo de exposición del tejido a la atmósfera. Un tiempo de coagulación riesgos a corto desprendimiento prematuro de las secciones de la MEA, mientras que la exposición prolongada a la atmósfera provoca degeneración de los tejidos. Porque la fuerza de la solución de trombina determina la velocidad del proceso de coagulación, es un parámetro muy importante para el éxito de unir las culturas saludables a la superficie de los AMUMA. Se obtienen mejores resultados con 1000 unidades (1KU; una unidad de NIH = 0.324 ± 0.073 mg). Es importante destacar que mezcla incompleta de los resultados de solución de plasma / trombina en la coagulación de la promoción de plasma espacial heterogénea-se rompe durante el cultivo. "Agujeros" en estos afectan gravemente a la salud plasma la cultura y separar la cultura de microelectrodos, comprometiendo así la calidad de los registros electrofisiológicos. Trabajar con placas de enfriamiento durante el MEA / tejido montaje disminuye la coagulación y permite la mezcla homogénea de la disolución de plasma / la trombina y el correcto posicionamiento de los cortes de tejido.
Del mismo modo, mediante la adición de medio de las gotitas individuales de la cámara de cultivo para sumergir la cultura después de tan sólo 5 minutos de la coagulación, reduce enormemente el riesgo de desprendimiento de los tejidos debido a las tensiones de la superficie. Una cultura de éxito se aplanan en la MEA y un poco ampliar mostrando un crecimiento saludable durante semanas, sin ningún signo importante de la incompleta del tejido y los AMUMA de contacto o la degeneración del tejido (por ejemplo, la figura. 1D). - Disección de los tejidos. Cuchillos Micro han mejorado mucho nuestro proceso de disección. Utilizamos dos hojas de afeitar borde (Herramientas de Bellas Ciencia - frágiles hojas de bisturí - 100050-00), de la cual nos separamos ~ 2 mm de ancho "hojas" con unos alicates, y los mantienen con el titular de bisturí. Las muestras de tejido disecado de la corte coronal con un movimiento suave, firme vertical de la hoja lo que reduce considerablemente el estrés mecánico debido a la tracción de los tejidos en general mejorar la salud de la cultura.
- Tejido de refrigeración. La correcta refrigeración de los cortes y secciones de tejido durante la preparación es esencial para el éxito de la cultura. Utilizamos a medida platos fríos construida de elementos Peltier adjunta debajo de un disco de metal. El calor producido por el elemento Peltier es eliminado a través de la perfusión de agua fría. Esto reduce considerablemente el tiempo de preparación y estandariza enfriamiento durante cada etapa de la preparación (Laboratorios Dold e Ingeniería 131 Plantación Dr. Seguin, TX 78155;. (830) 560 hasta 1471)).
- Condición de la incubadora. Una incubadora de medida con precisión las condiciones de oscilación interna ha sido crucial para nuestro éxito en el cultivo de segmentos en los acuerdos ambientales multilaterales. Basado en un original en la casa de diseño por Multichannelsystems (actualmente no disponible comercialmente), el dispositivo de oscilación interna consiste en bandejas que se unen a las dos ruedas laterales. Motores paso a paso y los controles computarizados permiten una trayectoria oscilante precisa (balanceo punto de vista, balanceándose velocidad y pausas intermitentes). En última instancia, las culturas corte deben ser expuestos al medio ambiente y la cultura en alternancia lento. El método tradicional consiste en colocar las culturas cortar en tubos estrechos que gire lentamente a lo largo de no más largo del eje. Aquí, la rotación lenta no produce estrés mecánico debido a la propia rotación y la velocidad de rotación es lo suficientemente alta para obtener una óptima "de alimentación / respiración" del ciclo de alrededor de 5 a 10 min de duración. El interior más compacto de la cámara de MEA, su volumen total de aproximadamente 2 cc, y el volumen medio de cultivo pequeña necesaria para el acondicionamiento medio por el propio tejido, plantea un reto importante. Por la oscilación entre la MEA ± ~ 70 º de ángulo (tiempo de ciclo: ~ 200 s), lo que frena la velocidad de balanceo, como las transiciones de la cultura entre la fase líquida y la atmósfera, y detener la oscilación en los ángulos extremos de una exposición prolongada a la atmósfera sido esencial para la supervivencia de la cultura.
2. Edad de desarrollo de las culturas corteza para estudiar las avalanchas neuronales
Rodajas aguda de la corteza de la rata se toma comúnmente a PND 0-1 y se cultivan durante varias semanas en el MEA. Los primeros estudios han demostrado claramente que una sola rebanada de culturas corteza, después de varias semanas in vitro, mantener una estructura en capas con los tipos de células de identificación que puede ser fácilmente comparado con las clases de células en vivo 18,18-21. La organización en capas en el sistema in vitro ha sido convenientemente utilizado para el estudio del tálamoinervación de la corteza durante el desarrollo de 22 a 24, así como para la conducción de las estructuras subcorticales como el estriado 25,26. De hecho, la especificidad en la formación de conexiones neuronales dentro de las regiones del cerebro permite la construcción de complejos sistemas in vitro de que la recuperación de numerosos sistemas de proyección detallada, por ejemplo, la de los ganglios basales, la corteza de circuitos 27-30.
Después de 4 - 6 semanas in vitro, de un solo rodajas de corteza 31 y rodajas de corteza co-cultivadas con estriado 26 o 32 tálamo muestran espontánea hacia arriba y hacia los estados se encuentran típicamente en vivo en ratas anestesiadas con uretano 33. La buena organización temporal de los estados arriba lleva el sello de anidado θ y γ-oscilaciones indicativo de una red electrofisiológica de las neuronas piramidales maduro y rápido remate interneuronas GABAérgicas 31. Es importante destacar que, en ausencia de dopamina D2-estimulación de los receptores, la maduración de parvalbúmina positivo interneuronas corticales se demora alrededor de 2 semanas en las culturas corte corteza 34. En línea con estos resultados, la evolución en el tiempo de desarrollo de anidado θ, β-y γ-se corresponde con las oscilaciones que en vivo cuando rebanadas corteza son co-cultivadas con el área tegmental ventral (VTA), que contiene las neuronas dopaminérgicas que se proyectan hacia el corteza 6.
Estos estudios indican que, al estudiar las avalanchas neuronales, que dependen fundamentalmente de la inhibición GABAérgica madura rápido y se encuentran en las capas superficiales de la corteza 2,4, el gran cuidado debe tenerse para asegurar la correcta maduración del tejido cortical. Mientras que las avalanchas neuronales surgen en las culturas corteza sola en el transcurso de tiempo de 2 - 5 semanas 4, cuando se requiere un curso de tiempo de desarrollo que se corresponde con el desarrollo en vivo, rodajas de corteza necesita un receptor adecuado de dopamina estimulación, por ejemplo, rodajas de corteza de cultivo co- con el VTA 6.
Disclosures
No hay conflictos de interés declarado.
Acknowledgments
Este estudio fue financiado por la División de Programa de Investigación Intramural (DIRP) del Instituto Nacional de Salud Mental, los Institutos Nacionales de Salud.
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}