

Summary
Une manière robuste à l'étude des avalanches neuronale, c'est à dire invariante d'échelle éclate l'activité spatio-temporelle, indicative de la dynamique de l'état critique dans le cortex. Avalanches émergent spontanément dans le développement de couches superficielles du cortex culture qui permet de mesures à long terme de l'activité avec des tableaux intégrés planaires multi-électrodes (MEA) dans des conditions contrôlées avec précision.
Abstract
Le cortex est spontanément actif, même en l'absence de toute entrée ou sortie particulière du moteur. Lors du développement, cette activité est importante pour la migration et la différenciation des types de cellules du cortex et de la formation des connexions neuronales 1. Dans l'animal adulte, l'activité continue reflète le passé et l'état actuel d'un animal dans lequel les stimuli sensoriels sont parfaitement intégrés pour calculer les actions futures. Ainsi, une compréhension claire de l'organisation de cours de l'activité spontanée savoir est une condition préalable pour comprendre la fonction du cortex.
De nombreuses techniques d'enregistrement a révélé que l'activité continue dans le cortex est constitué de nombreux neurones dont les activités individuelles transitoirement somme à des événements plus importants qui peuvent être détectés dans le potentiel de champ local (LFP) avec des microélectrodes extracellulaires, ou de l'électroencéphalogramme (EEG), le magnétoencéphalogramme (MEG ), et le signal BOLD de l'imagerie par résonance magnétique fonctionnelle (IRMf). La LFP est actuellement la méthode de choix lorsque l'on étudie l'activité de la population neuronale avec une résolution temporelle et spatiale à l'échelle mésoscopique (plusieurs milliers de neurones). Lors de la microélectrode extracellulaire, localement synchronisées activités de résultat neurones spatialement neighbored de déflexions rapides dans la LFP jusqu'à plusieurs centaines de microvolts. Lorsque vous utilisez un tableau de microélectrodes, les organisations de tels détournements peuvent être facilement surveillés dans l'espace et du temps.
Avalanches neuronale décrire l'organisation invariante d'échelle spatio-temporelle de l'activité neuronale dans le cerveau continue 2,3. Elles sont spécifiques à des couches superficielles du cortex comme établi in vitro, 4,5, in vivo chez le rat anesthésié 6, et chez le singe éveillé 7. Surtout, les études théoriques et empiriques suggèrent que les avalanches 2,8-10 neuronale montrent une dynamique bien équilibré état critique du cortex qui optimise le transfert de l'information et de traitement de l'information.
Afin d'étudier les mécanismes du développement neuronal avalanches, l'entretien et la réglementation, dans des préparations in vitro sont très bénéfiques, car ils permettent des enregistrements d'activité stable avalanches dans des conditions contrôlées avec précision. L'actuel protocole décrit comment étudier les avalanches neuronales in vitro, en profitant du développement de la couche superficielle du cortex dans les cultures organotypiques, les cultures tranche-dire, cultivé sur planaires, les tableaux de microélectrodes intégré (AME; voir aussi 11-14).
Protocol
1. Stérile, la Chambre de verre plombable avec la MEA pour les enregistrements à long terme
- Cylindres en verre fileté avec Téflon capuchon plastique (Ace Glass), nécessaires à la clôture de la culture de chambre sécurisée et serrés, sont coupés (Aceglass) d'environ 2 mm du fond du filet (Fig. 1A, B). Anneaux de verre propre par un rinçage à l'eau (3x) et l'ébullition pendant 5 min dans 200 la preuve alcool éthylique, laisser sécher.
- Solution de silicone Aliquot nécessaire pour attacher les anneaux de verre à la surface de la MEA. Mélangez 15 ml de parties A & B de Sylgard 184 Kit élastomère de silicone complètement, laisser reposer pendant 15 min pour éliminer les bulles d'air, de stocker en aliquotes de 1 ml à -20 ° C.
- Anneau de verre Colle à l'AEM (8x8 grille w / électrode de masse interne, diamètre de l'électrode 30 um, um 200/100 distance inter-électrodes pour le rat / souris) (Fig. 1A, B). Prélever 1 ml de silicium (23 ° C) dans une seringue avec une aiguille de petit calibre. Appliquez de la silicone à la surface de la coupe du mat anneau de verre, anneau de verre du centre sur la MEA, appliquer une couche supplémentaire de silicone autour de l'extérieur de l'anneau d'une forte étanchéité, laisser sécher pendant 1 - 2 heures à ~ 60 ° C sur une plaque chaude.
- Stériliser MEA chambre et bouchons de chambre dans une hotte à flux laminaire par rincer à l'eau déminéralisée 3x suivi par l'alcool à 70% (3 x; pour dernier rinçage laisser reposer pendant 10 min dans de l'alcool), suivies de 10 min d'exposition de la chambre et l'intérieur de la PAC à la lumière UV . Autoclave MEA chambre (120 ° C; humide; min 45) et laisser sécher.
- Manteau MEA surface intérieure chambre de culture avec de la poly-D-lysine. Pour de nouveaux AME, qui sont plutôt lipophiles, manteau par aspiration de gouttelettes répétées de la solution de la grille d'électrodes. Pour AME utilisé, couvrir le fond avec une solution de chambre, aspirer l'excès de liquide, laisser évaporer dans des conditions stériles à l'intérieur capuche laminaire. Attacher bouchon pour sceller la chambre MEA pour le stockage et l'utilisation future.
2. Ingrédients nécessaires à la préparation et la croissance des cultures organotypiques
- Dissoudre la gélose stérile de NaCl 0,9%, versez-le dans boîte de Petri stérile (Falcon, 100 x15; niveau de ~ 5 mm), laisser refroidir et emballage stérile avec du Parafilm pour le stockage. Couper 20 x 10 x 5 mm à partir de blocs de gélose solide pour l'utilisation.
- Super Store colle, par exemple Devcon Super Glue II, à l'emballage essuyées avec EtOH 70% avant l'ouverture, à l'intérieur de la hotte à flux laminaire pour préserver la stérilité.
- Préparer 50% de D-glucose (SIGMA ultra, G7528) en ajoutant 40 g de glucose à 40 ml d'eau déminéralisée de la culture (Sigma). Conserver dans aliquots de 2 ml à -20 ° C.
- Ajouter 4 ml de 50% de D-glucose pour 500 ml de solution saline équilibrée de Gey, et réfrigérer au gadoue (mélange de liquide / cristaux de glace) dans le congélateur avant de les utiliser.
- Plasmatique de poulet reconstituer dans 5 ml d'eau déminéralisée de la culture (secouer doucement, éviter la formation de bulles), laissez reposer la solution pendant 5 - 10 min, agiter doucement, et le contenu décantat claire dans un boîte de Pétri stérile. Magasin de solutions plasma, partie aliquote 350 pi dans des cryotubes (NuncTM), à -20 ° C; Stérile-filtre (filtre de 0,22 um protéines des pores)
- Reconstituer la thrombine du plasma bovin en conséquence, stérile filtre (filtre de 0,22 um de porosité), 40 pl aliquote dans des cryotubes (NuncTM), conserver à -20 ° C. Pour la solution de travail, diluer 40 pl de la solution de thrombine à 375 ul de la solution saline équilibrée de Gey w / D-glucose.
- Préparer 400 ml de milieu de culture en mélangeant 100 ml de sérum de cheval, 200 ml de milieu basal Eagle, 100 ml de Hank solution saline tamponnée à laquelle 4 ml de glucose à 50% et 2 ml de 200 mM de L-glutamine sont ajoutés. Peut être conservé 4 - 8 semaines dans 100 ml bouteilles PYREX à 4 ° C.
- Préparer inhibiteur de la mitose en mélangeant 0,3 mM d'uridine, 0,3 mm mm ARA-C-β-cytosine D-arabinofuranoside, et 0,3 de 5-fluoro-2'-désoxyuridine, filtre stérile, partie aliquote de 200 ul et conserver à -20 ° C pour les 6 - 12 mois.
3. Cortex et ventrale tegmentale dissection des tissus Zone (VTA) (durée: <1h)
- Procédure rendements cortex et des coupes de tissus VTA pour ~ 12 co-cultures à partir de rats ou de souris, et elle est disposée à l'intérieur d'une hotte à flux laminaire dans des conditions stériles. Le temps total pour la collecte de tissus devrait être de 1 hr.
- Prenez en bonne santé, bien nourris chiots (taille de la portée ~ 10; présence d'un «spot lait» abdominale) à 1 - 2 jours postnatale (DPN). Tenir un chiot doucement par le nez, lui permettent de pendre librement, rapidement et de décapiter à la base du cou avec des ciseaux pointus.
- Pour le retrait du cerveau, enlever la peau (deux latérales coupes ciseaux), découper le crâne ouvert avec ciseaux bel œil (1 sagittales médianes coupées; 1 coupe coronale au cortex / jonction du cervelet). Retournez en arrière tous les quatre volets du crâne. Avec une spatule aiguisé, couper frontalement à travers le bulbe olfactif, une spatule d'avance caudalement au-dessous du cerveau. Soulevez doucement le cerveau du crâne et laissez-le glisser dans stérile, réfrigérée, la solution de Gey pour le refroidissement rapide et un stockage temporaire. Répétez les étapes de 3,2 à 3,3 pour les deux cerveaux de plus (durée totale: <20 min).
- Pour obtenir VTAtissus, le transfert des cerveaux sur une stérile, boîte de Petri à sec en utilisant une petite spatule. Continuer à supprimer les excès de liquide en glissant doucement chaque cerveau d'environ 1 cm de côté. Retirer du tronc cérébral par une coronale, coupe verticale au niveau du cervelet en utilisant une lame de rasoir.
- Bloc de gélose colle sur le montage de disque pour la stabilisation mécanique du cerveau lors de tranchage procédure. Placez une fine ligne de colle de quelques millimètres à l'avant du bloc d'agar sur le disque (éviter la colle de toucher la gélose).
- En utilisant une petite spatule, transférer et monter chaque cerveau, pôle frontal vers le bas. Assurez-vous que les pôles frontaux sont collés sur le disque de montage et que les côtés ventrale toucher la gélose sans aucun résidu de colle afin de parvenir à la stabilisation mécanique correcte lors de la coupe et facile d'envol des tranches coupées.
- Soigneusement submerger et sécuriser le disque de montage avec l'assemblage du cerveau dans un bac vibratome (par exemple, Leica VT1000) rempli de solution stérile, réfrigérée Gey. Avec une lame de rasoir soigneusement nettoyés (90% EtOH), couper des tranches coronales du mésencéphale à une fréquence plus élevée de vibration et relativement faible vitesse de marche avant à une épaisseur de 400 500 um. En utilisant une pipette Pasteur inversé avec une ampoule d'aspiration, de transfert et de recueillir les tranches contenant l'aire tegmentale ventrale en 35 x 10 mm boîte de Pétri remplie de solution stérile, réfrigérée Gey (Fig. 1C; voir la plaque a également coronales de 18 à 20 au E22 en 15).
- Pour les sections du cortex, répétez les étapes 3.2 à 3.6, mais s'appliquent coupe verticale entre le cortex et le cervelet, et monter avec le pôle frontal forebrains place. Environ 3 tranches coronales (350 um d'épaisseur) à partir du niveau du striatum sont collectées pour la dissection du cortex avenir.
- Avec un couteau micro-faite de lames de rasoir cassée, disséquer ~ 2 mm de large section coronale cortex frontal et les zones du mésencéphale contenant l'aire tegmentale ventrale (Fig. 1C), sous un stéréomicroscope. Recueillir des coupes de tissus séparément dans des petits plats (diapositives de chambre, par exemple) rempli d'une solution de Gey réfrigérée.
4. Montage Cortex et des tranches de tissu VTA sur le MEA (Heure: <1 h)
- Position MEA à température ambiante sous un microscope stéréo avec le réseau d'électrodes dans le foyer. Centre d'une goutte 25 pl de plasma sur le propre, sans poussière, et la matrice stérile réseau d'électrodes. Utiliser spatules petit, glissez soigneusement un cortex et la section VTA dans la goutte de plasma.
- Placer sur une plaque de refroidissement MEA, recentrer vue, laissez refroidir pendant environ 15 s, puis ajouter 25 ul de la thrombine dans le plasma de gouttelettes. Avec la pointe de thrombine pipette, soigneusement étaler le mélange plasma / thrombine avec de petits mouvements circulaires à travers la MEA. Ne touchez pas le réseau d'électrodes cassantes directement. Doucement la position du cortex sur le tableau avec bordure dorsale le long de la ligne la seconde électrode de la matrice. De cette façon, les couches superficielles de développement portera éventuellement sur le rappel du tableau. Le VTA est placé à côté de la frontière ventrale de la section du cortex (Fig. 1D).
- Cap et lâchement fermer la chambre MEA de conserver une humidité élevée tandis que l'assemblage MEA / culture s'assoit pour ~ 5 min à l'intérieur du capot à la température ambiante pour permettre plasma / thrombine coagulation. Pendant ce temps, répétez l'étape 4.1 à 4.3 pour les trois cultures plus.
- Ajouter avec précaution 600 pi de milieu de culture dans de petites gouttelettes dans la chambre de culture en utilisant une seringue de 1 cc avec 25 x 5 / 8 aiguille.
- Fermer hermétiquement la chambre AME et le lieu MEA / montage de la culture sur le plateau de rangement à bascule intérieur de l'incubateur (Fig. 1B). Pour accélérer la procédure, 3 - 4 AME peuvent être assemblés dans des séquences qui se chevauchent. Temps de montage pour 12 AEM devrait être <1hr.
- Après 2 jours in vitro (DIV), ajouter 10 ul d'inhibiteur de la mitose. Actualiser la culture des médias de 60% à 4 DIV et tous les 4 jours après.
5. Enregistrement électrophysiologique et la génération de stimuli
- Pour établir la relation entre les déformations importantes dans le potentiel de champ local (LFP) et la tendance des neurones au feu des potentiels d'action, après une activité record ~ 5,6 semaines spontanée à 24 kHz pour ~ 10 min de chaque électrode du MEA (Hardware : MEA1060 w / circuit de découpage, x1200 gain, 12 bits A / D, la gamme 0-4096 mV, systèmes multicanaux; Logiciel: MC_Rack). Au sol est assurée soit par l'électrode de terre interne ou en externe par l'ajout d'une cellule Ag / AgCl-moitié.
- Séparée de la LFP avec un filtre passe-bande de 1 - 200 Hz du pic d'activité extracellulaire (300 passe-bande - 3000 Hz). L'activité de Spike peuvent être classées en activité unique et multi-appareil à l'aide hors ligne trieurs pic (par exemple Plexon Inc.) Calculer pic déclenché moyennes pour chaque électrode. Pour les cultures de cortex, la plupart des moyennes identifiera négative déflexions LFP (PFNL) comme le temps privilégié du dopage neuronale dans la culture.
- Calculer pour chaque électrode d'un seuil de -3 écarts types du bruit (DS) à partir des traces LFP (Fig. 2), Déterminer les périodes de pointe et les amplitudes des nLFPs ce seuil croix (Fig. 2B, C). Choisissez un bac de temps At (par exemple entre 2 - 8 ms) et d'identifier les pôles de PFNL spatio-temporelle sur le tableau par la concaténation de tous nLFPs électrode qui sont dans la même combine de temps successifs de At longueur (Fig. 2D; pour plus de détails voir le 2,4 , 5).
- Afin d'identifier les avalanches neuronale, calculer la taille de chaque cluster PFNL, par exemple le nombre d'électrodes actives ou la somme des amplitudes PFNL, construire un histogramme de taille, et l'intrigue en double-logarithmique coordonnées. Pour les avalanches neuronale, la distribution de la taille suit une loi de puissance approchée par une ligne droite en double-logarithmique coordonnées 2 (Fig. 2E, F). Voir 16 pour les tests statistiques sur les lois de puissance.
- Susciter des réponses évoquées dans le tissu en choisissant une électrode à travers lequel le courant contrôlé par des stimuli avec une amplitude S sont appliqués (générateur de stimulation STG 1008, systèmes multicanaux). Pour réduire les dommages électrode, utiliser une gamme limitée, chargé neutre stimulation des chocs simples bipolaire carrés de forme d'onde: 50 ms avec une amplitude-S suivie par 100 us avec une amplitude + / S 2 et S entre 10 - 200 uA. Voir le manuel du propriétaire pour plus de détails.
- Pour enregistrer la gamme dynamique 9, relance records enregistrés réponses LFP à 4 kHz fréquence d'échantillonnage sur toutes les électrodes suivantes 500 ms après la stimulation. Utilisez obturation des circuits (systèmes multi-canaux) qui déconnecte les amplificateurs stade de la tête lors de la stimulation pour réduire les artefacts de stimulation et de prévenir la pré-amplificateur de saturation.
6. Les résultats représentatifs:
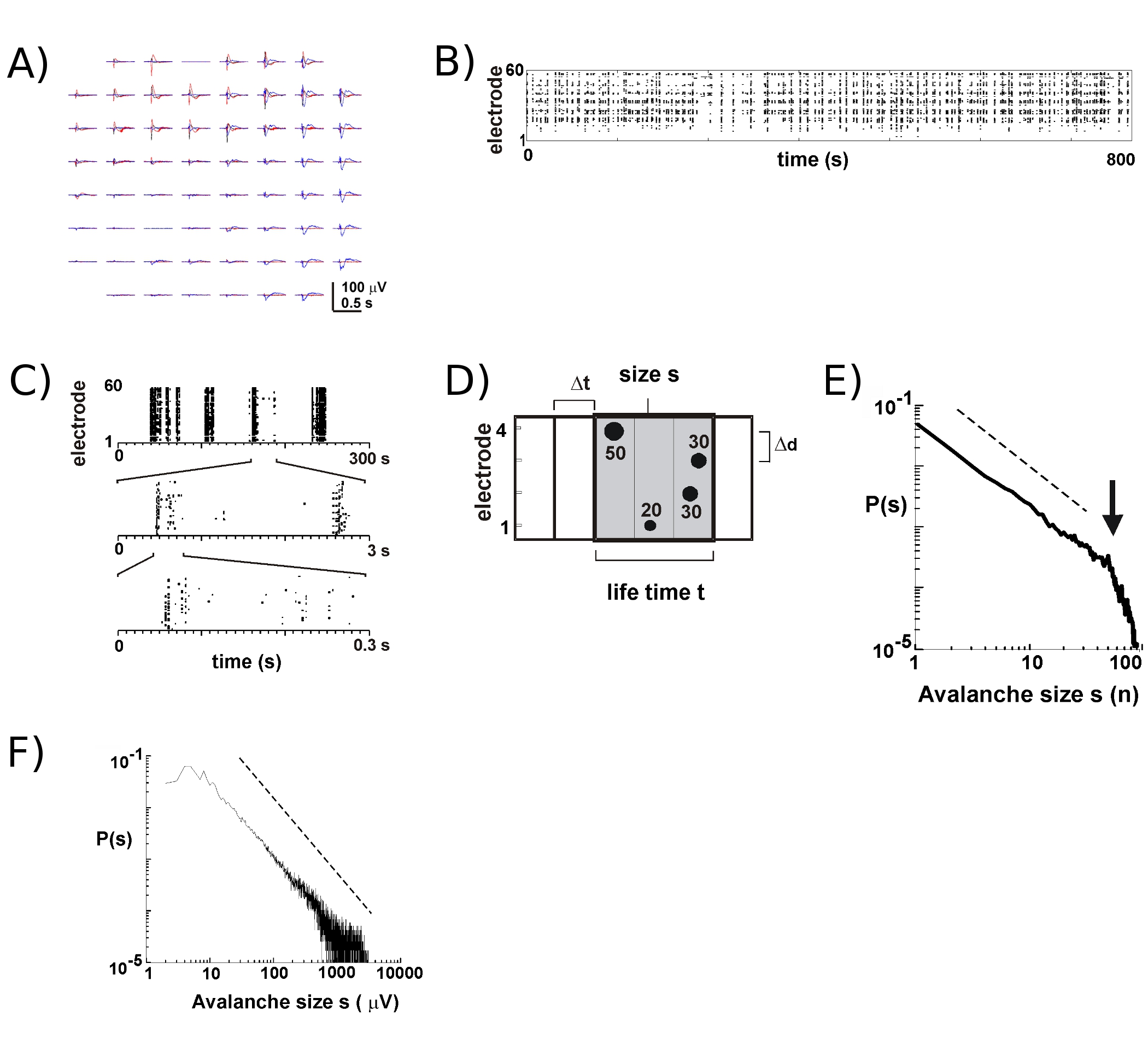
Avec de nouveaux AME environ 8 - 9 sur 10 cultures vont survivre pendant plusieurs semaines. La plupart de nos enregistrements à long terme aura lieu à l'intérieur de l'incubateur en milieu de culture, ce qui nous permet de suivre le développement des cultures individuelles au cours de plusieurs semaines 5. Basé sur nos expériences, les enregistrements LFP peut être obtenu de façon fiable avec les AME utilisé pour plus de 100 jours de culture. En revanche, l'activité extracellulaire pic est plus mesurée de manière fiable avec relativement nouveaux AME (<40 jours de culture). Dans une expérience typique, nous transférons un AME dans le bac de stockage (Fig. 1B, à droite) pour le bac avec l'étape de tête attachée (Fig. 1B, à gauche) en maintenant la chambre de culture hermétiquement fermés. Pour le cortex 5, le cortex-VTA co-cultures 6, ainsi que chez le rat anesthésié 6 et le singe éveillé in vivo 7, décharges neuronales au cours des avalanches neuronale dans les couches superficielles se produit principalement près de la déviation de crête négative de la LFP (PFNL ). Ainsi, l'organisation spatio-temporelle des locaux synchronisés groupes neuronaux peuvent être estimées en mesurant l'apparition de nLFPs dans l'espace et le temps sur les 17 tableaux.
L'activité sur le MEA tend à émerger dans les grappes temporelles, telles que l'activité moins une électrode est accompagné d'une activité à d'autres sites. Formes d'onde typique de la LFP lors de périodes d'activité tels sont présentés dans la figure 2A, par plus de 3 pôles de comploter survenant quelques secondes d'intervalle. Pour chaque cluster, déflexions champ négatif peut être vu à plusieurs électrodes dans une fenêtre de 1 s. Lors de l'extraction des pics PFNL qui traversent un seuil de multiples négative SD, l'activité sous la forme de PFNL heures de pointe est idéalement visualisé dans une trame dans laquelle «colonnes» de points représentent près nLFPs coïncidant au différentes électrodes (figure 2B). L'organisation spatio-temporelle de cette activité est assez complexe; «colonnes», qui apparaissent plus ou moins homogène au faible résolution temporelle, sont composées de grappes séparées à résolution temporelle plus élevée et ainsi de suite (figure 2C). En fait, l'émergence de clusters PFNL spatio-temporelle est très organisé dans les réseaux corticaux. Plus précisément, l'organisation est invariante d'échelle pour les avalanches neuronale. Cela est démontré par le calcul de la probabilité de tailles de cluster à un At résolution temporelle donnée. Ici, les grappes sont composées de nLFPs qui se produisent dans les bacs en même temps ou successivement (Fig. 2D). Lorsque la taille d'un tel pôle est exprimé en nombre total de nLFPs par cluster, ou intégré amplitudes PFNL par cluster, les distributions de taille des clusters révèle une loi de puissance, dont la pente a été montré pour être 2,4,5,7 -1,5 ( Fig. 2E, F). Notez que cette distribution identifie une commande invariante d'échelle des tailles de cluster qui est le rapport des tailles S à KXS, où k est un facteur constant, est de -1,5 K, qui est indépendant de s. Cette organisation en loi de puissance est indépendante de la taille du tableau 2, la résolution temporelle At 2, et le seuil utilisé pour identifier PFNL significative déflexions 7. Parce que les échelles d'amplitude PFNL avec la taille du groupe 7 neuronale, l'organisation invariante d'échelle des nLFPs reflète une invariante d'échelle, c'est à dire fractale, ordonnant of localement synchronisées groupes de neurones qui incluent toutes les tailles.

La figure 1 (A). Vues latérales et le dessus de la MEA avec anneau de verre fileté monté, et le bouchon correspondant. (B) Vue de l'intérieur de l'incubateur. Gauche: headstage montage permettant d'enregistrer à partir d'une culture unique sous condition incubateur. Droite: Bac tenant AME nombreuses à la croissance de la culture. Côté roues: moteur pas à pas l'appareil commandé à bascule pour alternant phase de plongée et son atmosphère exposés nécessaires à la croissance de la culture. Schéma (C) pour les tranches coronales rats utilisés pour Cortex-VTA co-cultures. Sections Cortex (à gauche) et les articles du mésencéphale (au milieu, à droite) contenant le VTA aire tegmentale ventrale (VTA; gris) sont obtenues par découpe le long des lignes brisées. CTX: cortex; WM: la substance blanche; CPU: striatum; VTA: Pons: zone pontique. Voir aussi les plaques correspondantes coronale 8, 18, et 20 par 15. (D) Le placement et la croissance d'un seul cortex VTA co-cultures de la MEA et son développement au cours des 9 premiers DIV dans la culture. Notez l'aplatissement de la culture et de son expansion progressive sur le tableau. Pièces de tissu réfléchissant indiquent les cellules dégénérées et les débris tissulaires. Les tissus sains est opaque et grisâtre sous transillumination avec la lumière visible.

Figure 2. Avalanches neuronale du cortex cultures organotypiques. (A) de trois Overplot période d'activité spontanée sur le tableau, séparés par quelques secondes. Notez que chaque période d'activité se compose de déflexions LFP négatif sur de nombreuses électrodes sur le tableau (chaque couleur d'étiquettes une période d'activité). (B) les périodes de pointe négative de nLFPs de chaque électrode sont assemblés dans une trame d'activité. «Column' structures semblables ont indiqué des périodes d'activité synchrone proche. (C) Notez que les colonnes qui apparaissent hautement synchronisée à une échelle de temps se composent de plusieurs colonnes à l'échelle temporelle plus élevée (3 échelles temporelles montré). Représentation schématique (D) de l'algorithme d'avalanche neuronale. Sur une 2 x 2 heures de pointe d'électrode tableau et l'amplitude des débattements LFP négatif (PFNL) de franchir un seuil de x-SD du bruit sont identifiés. L'organisation spatio-temporelle de nLFPs est regroupé dans des bacs de temps successivement active de At largeur. La taille d'un cluster est identifié soit par le nombre de sites actifs, c'est à dire avec des électrodes PFNL (s = 4), ou la somme des amplitudes intégrées PFNL (s = 130 mV). La durée de vie est mesurée en multiples de AT. Loi de puissance (E, F) dans la distribution de taille de cluster identifie les clusters que les avalanches neuronale. Notez que le choix d'une distance inter-électrodes notamment Ad pour le tableau (ici 200 um) introduit un At notamment au cours de laquelle la dynamique doit être observé. Plus précisément, le rapport dans lequel Ad / At rapproche de la moyenne de vitesse de propagation dans le réseau, à laquelle la pente α de la loi de puissance est approximativement -1,5 pour les avalanches neuronale 2,4,5. S'il vous plaît cliquez ici pour voir une version agrandie de la figure 2.
Discussion
1. Problèmes techniques:
- Technique stérile. La préparation de l'AME et la préparation de la culture sont toutes effectuées dans une hotte à flux laminaire dans des conditions stériles. Les antibiotiques, qui affectent l'activité neuronale, ne sont pas utilisées à tout moment pendant le processus de préparation et de la culture.
- Plasma / thrombine coagulation et l'adhérence des tissus sur la survie des AEM. Tissu sur la MEA exige un équilibre délicat entre le temps requis pour le plasma / thrombine coagulation et le temps d'exposition des tissus à l'atmosphère. Un temps de coagulation courte risques décollement prématuré des sections de la MEA, tandis que l'exposition prolongée à l'atmosphère provoque la dégénérescence des tissus. Parce que la force de la solution de thrombine détermine la vitesse du processus de la coagulation, il est un paramètre très important pour réussir à fixer, des cultures saines à la surface de la MEA. Nous obtenons de meilleurs résultats à l'aide de 1000 unités (1KU; 1 unité = 0,324 ± NIH 0,073 mg). Surtout, mélange incomplet des résultats solution de plasma / thrombine dans la coagulation du plasma spatialement hétérogènes promotion-ruptures en cours de culture. «Trous» Ces plasmatique gravement affecter la santé de la culture et de détacher la culture de microélectrodes, compromettant ainsi la qualité de l'enregistrement électrophysiologiques. Travailler avec des plaques de refroidissement lors de la MEA / tissu montage ralentit la coagulation et permet un mélange homogène de la solution de plasma / thrombine et le bon positionnement des sections de tissus.
De même, en ajoutant le milieu dans des gouttelettes unique pour la chambre de culture de submerger la culture, après seulement 5 min de la coagulation, réduit considérablement le risque de détachements de tissu due à des tensions de surface. Une culture réussie va aplatir sur la MEA et légèrement expansion montrant une croissance saine pendant des semaines, sans aucun des signes importants de tissus incomplète-MEA contact ou la dégénérescence des tissus (par exemple, Fig. 1D). - Dissection des tissus. Couteaux Micro ont grandement amélioré notre processus de dissection. Nous utilisons des lames de rasoir à double pointe (Outils Fine Science - cassables les lames de bistouri - 100050-00), à partir de laquelle nous avons divisé ~ 2 mm de large "lames" en utilisant une pince, et les maintenir avec un support de scalpel. Des échantillons de tissus disséqués de la tranche coronale en utilisant une surface lisse, mouvement ferme verticale de la lame ce qui réduit considérablement les contraintes mécaniques dues à la traction du tissu améliorer la santé globale de la culture.
- Refroidissement des tissus. Refroidissement correct des tranches et des coupes de tissus lors de la préparation est essentielle pour la réussite de la culture. Nous utilisons des faits sur des assiettes froides construites d'éléments Peltier attaché en dessous pour un disque de métal. La chaleur produite par l'élément Peltier est enlevé par une perfusion d'eau froide. Ceci réduit considérablement le temps de préparation et standardise le refroidissement pendant chaque étape de la préparation (Labs Dold & Engineering 131 Plantation Dr Seguin, TX 78155;. (830) 560-1471)).
- Incubateur état. Un incubateur sur mesure avec précision les conditions de bascule interne a été essentielle à notre succès à cultiver les tranches sur les AEM. Basé sur un original en interne la conception par Multichannelsystems (actuellement non disponible dans le commerce), l'appareil bascule interne est composé de plateaux qui sont attachés aux deux roues grand côté. Les moteurs pas à pas et des contrôles informatisés permettant une trajectoire précise à bascule (rocking angle, bascule la vitesse, et pauses intermittentes). Finalement, les cultures tranche besoin d'être exposés à moyenne atmosphère et la culture en alternance lente. L'approche traditionnelle est de placer les cultures tranche dans des tubes étroits qui tournent lentement le long il axe le plus long. Ici, la rotation lente ne produit pas de contraintes mécaniques dues à la rotation elle-même, et la vitesse de rotation est suffisamment élevée pour obtenir un optimal »d'alimentation / respiration» du cycle d'environ 5 à 10 min durée. L'intérieur plus compact de la chambre de la MEA, son volume total d'environ 2 cc, et le volume de culture petit moyen nécessaire pour le conditionnement de support par le tissu lui-même, représente un défi important. En basculant la MEA entre ± ~ 70 ° d'angle (temps de cycle: ~ 200 s), ce qui ralentit la vitesse de balancement que les transitions de la culture entre la phase liquide et l'atmosphère, et l'arrêt de la bascule au niveau des angles extrêmes d'une exposition prolongée à l'atmosphère a été essentielle pour la survie de la culture.
2. Le stade de développement des cultures à l'étude du cortex avalanches neuronale
Tranches de cortex de rats aiguë sont souvent prises au PND 0-1 et cultivées pendant plusieurs semaines sur la MEA. Les premières études ont clairement démontré que seule tranche de cultures cortex, après plusieurs semaines in vitro, de maintenir une structure en couches avec différents types cellulaires identifiables qui peuvent être facilement comparées aux classes de cellules in vivo 18,18-21. L'organisation en couches dans ce système in vitro a été utilisée pour étudier commodément thalamiqueinnervation du cortex pendant le développement 22-24, ainsi que pour la conduite des structures sous-corticales comme le striatum 25,26. En fait, la spécificité de la formation des connexions neuronales au sein et entre les régions du cerveau permet la construction de complexes systèmes in vitro que la récupération de nombreux systèmes de projection détaillée, par exemple, que des ganglions de la base du cortex-circuits 27-30.
Après 4 - 6 semaines in vitro, seule tranches et des tranches de cortex 31 cortex co-cultivées avec striatum 26 ou 32 montrent thalamus spontanée haut et le bas on trouve habituellement dans les États vivo chez le rat anesthésié uréthane 33. L'organisation temporelle fine de ces états-place porte le poinçon de imbriquées θ et γ-oscillations indicative d'un réseau électrophysiologique des neurones pyramidaux matures et fast-dopage interneurones GABAergiques 31. Surtout, en l'absence de dopamine récepteur D2 de stimulation, la maturation de la parvalbumine-positif interneurones corticaux est retardée d'environ deux semaines dans les cultures tranche de cortex 34. En ligne avec ces résultats, le cours du temps de développement de imbriquées θ-, β-et γ-oscillations est adaptée à celle in vivo lorsque tranches cortex sont co-cultivées avec l'aire tegmentale ventrale (VTA), qui contient les neurones dopaminergiques se projetant vers l' corticale 6.
Ces études indiquent que lorsque l'on étudie les avalanches neuronale, qui dépendent essentiellement matures inhibition GABAergique rapide et sont situés dans les couches superficielles du cortex 2,4, le plus grand soin doit être pris pour assurer la maturation adéquate du tissu cortical. Alors que les avalanches surviennent dans les cultures neuronales du cortex seule au cours du temps de 2 - 5 semaines 4, en exigeant un cours à temps de développement qui est adaptée à la développement in vivo, des tranches de cortex ont besoin de stimulation d'un récepteur approprié de la dopamine, par exemple, par tranches de cortex co-culture avec le VTA 6.
Disclosures
Aucun conflit d'intérêt déclaré.
Acknowledgments
Cette étude a été financée par la Division du programme de recherche intra-muros (PRID) de l'Institut national de santé mentale, National Institutes of Health.
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}