

Summary
A forma robusta para estudar avalanches neuronal, ou seja, escala-invariante rajadas espaço-temporal atividade, indicativo da dinâmica do estado crítico no córtex. Avalanches surgem espontaneamente no desenvolvimento de camadas superficiais do córtex cultura que permite medições a longo prazo da atividade com arrays planar integrada multi-eletrodo (MEA), sob condições estritamente controladas.
Abstract
O córtex é espontaneamente ativa, mesmo na ausência de qualquer determinada entrada ou saída do motor. Durante o desenvolvimento, essa atividade é importante para a migração e diferenciação de tipos de células do córtex e da formação de conexões neuronais 1. No animal adulto, a atividade em curso reflete o passado eo presente estado de um animal em que os estímulos sensoriais estão perfeitamente integrados para calcular as ações futuras. Assim, uma compreensão clara da organização da atividade em curso ou seja espontânea é um pré-requisito para entender a função do córtex.
Técnicas de gravação numerosos revelaram que a atividade em curso no córtex é composto de muitos neurônios individuais cujas atividades transitoriamente soma para eventos maiores, que podem ser detectados no potencial de campo local (LFP), com microeletrodos extracelulares, ou no eletroencefalograma (EEG), o magnetoencephalogram (MEG ), eo sinal BOLD da ressonância magnética funcional (fMRI). A LFP é atualmente o método de escolha quando estudar a atividade neuronal população com resolução temporal e espacial de alta na escala mesoscópica (vários milhares de neurônios). No microeletrodos extracelulares, localmente sincronizado atividades espacialmente neighbored resultado neurônios em deflexões rápido no LFP até várias centenas de microvolts. Quando usando uma matriz de microeletrodos, as organizações de tais desvios podem ser convenientemente monitoradas no espaço e no tempo.
Avalanches neuronal descrever a organização em escala espaço-temporal invariante da atividade neuronal em curso no cérebro 2,3. Eles são específicos para as camadas superficiais do córtex, conforme estabelecido in vitro 4,5, in vivo no rato anestesiado 6, e no macaco acordado 7. Importante, estudos teóricos e empíricos sugerem que 2,8-10 avalanches neuronal indicar uma dinâmica perfeitamente equilibrada estado crítico do córtex que otimiza a transferência de informações e processamento de informações.
A fim de estudar os mecanismos de desenvolvimento avalanche neuronal, manutenção e regulamentação, em preparações in vitro são altamente benéficos, uma vez que permitem gravações de actividade estável avalanche em condições estritamente controladas. O protocolo atual descreve como estudar avalanches neuronal in vitro, aproveitando o desenvolvimento da camada superficial em culturas organotípicas córtex, culturas fatia isto é, cultivadas em planar, matrizes de microeletrodos integrados (MEA, ver também 11-14).
Protocol
1. Estéril, com câmara de vidro Sealable MEA para gravações a longo prazo
- Cilindros de vidro com rosca e tampa de plástico com Teflon (Ace Glass), necessárias para o fechamento seguro e apertado cultura de câmara, são cortados (Aceglass) cerca de 2 mm da parte inferior do fio (Fig. 1A, B). Anéis de vidro limpo com agua (3x) e fervente por 5 min em 200 provas álcool etílico, deixe secar.
- Solução de silício alíquota obrigado a juntar anéis de vidro à superfície MEA. Misture 15 ml de peças de A & B de Sylgard Kit de elastômero de silicone 184 completamente, deixe descansar por 15 min para remover bolhas de ar, uma loja em alíquotas ml a -20 ° C.
- Cola anel de vidro para o MEA (8x8 grade w / eletrodo de aterramento interno, 30 diâmetro do eletrodo M, 200/100 M-eletrodo inter distância para rato / mouse) (Fig. 1A, B). Tome-se 1 ml de silicone (23 ° C) em seringa com agulha de calibre pequeno. Aplique silicone para superfície de corte do anel de vidro polido, anel de vidro no centro MEA, aplique uma camada adicional de silício em torno do exterior do anel de vedação para uma mais forte, vamos cura para 1-2 horas a ~ 60 ° C em chapa quente.
- Esterilizar MEA câmara e as tampas de câmara em uma capela de fluxo laminar por lavar 3x em água deionizada seguido pelo álcool a 70% (3 x; para o último enxágüe deixe descansar por 10 min em álcool), seguido de 10 min de exposição da câmara e tampa interior à luz UV . Autoclave MEA câmara (120 ° C; molhado; 45 min) e deixe secar.
- Revestimento de superfície MEA dentro da câmara de cultura com poli-D-lisina. MEAs para novas, que são bastante lipofílico, casaco por aspiração de gotículas repetidas da solução da grade do eletrodo. Para MEAs usado, tampa inferior da câmara com a solução, aspire o excesso de líquido, deixe evaporar sob condições estéreis dentro capô laminar. Anexar tampa para selar MEA câmara para armazenamento e uso futuro.
2. Ingredientes necessários para a preparação e Crescimento das Culturas organotípicas
- Dissolver ágar estéril em 0,9% NaCl, despeje em placa de Petri estéril (Falcon, 100 x15; ~ nível 5 mm), deixe esfriar e enrole estéril com Parafilm para armazenamento. Corte de 20 x 10 x 5 mm a partir de blocos de agar sólido para uso.
- Loja super cola, por exemplo, Devcon Super Glue II, com a embalagem varrida para baixo com EtOH 70% antes da abertura, dentro da capela de fluxo laminar para preservar a esterilidade.
- Prepare 50% D-Glicose (SIGMA ultra, G7528), adicionando 40 g de glicose a 40 ml de água desionizada cultura (Sigma). Armazenar em alíquotas de 2 ml a -20 ° C.
- Adicionar 4 ml de 50% D-glicose por 500 ml de solução salina balanceada de Gey, e frio de lama (mistura de líquido / cristais de gelo) no congelador antes de usar.
- Reconstituir plasma de frango em 5 ml de água da cultura de-ionizado (agitação suave, evitar a formação de bolhas), deixe a solução de se contentar com 5 - 10 min, rode suavemente, e decantate o conteúdo claro em uma placa de Petri estéril. Armazenar a solução plasma, 350 mL da alíquota em criotubos (NuncTM), a -20 ° C.; estéril-filtro (proteína 0,22 mM poros do filtro)
- Trombina reconstituir a partir de plasma bovino em conformidade, estéril-filtro (0,22 mM poros do filtro), alíquota de 40 mL em criotubos (NuncTM), armazenar a -20 ° C. Para trabalhar solução, diluir 40 mL da solução de trombina em 375 ml de solução salina balanceada de Gey w / D-Glicose.
- Prepare 400 ml de meio de cultura através da mistura de 100 ml de soro de cavalo, 200 ml Basal Medium Eagle, 100 ml da solução de Hank Buffered Saline a que 4 ml de glicose 50% e 2 ml de 200 mM L-glutamina são adicionados. Podem ser armazenados 4-8 semanas, em frascos de 100 ml PYREX a 4 ° C.
- Prepare inibidor da mitose através da mistura de 0,3 mM uridina, 0,3 mM mM ARA-C citosina-β-D-arabinofuranoside e 0.3 5-fluoro-2'-deoxiuridina, filtro estéril, 200 mL da alíquota e armazenar a -20 ° C por 6 - 12 meses.
3. Córtex e área tegmental ventral dissecção dos tecidos (VTA) (hora: <1h)
- Procedimento de rendimentos córtex e cortes de tecidos VTA para ~ 12 co-culturas de ratos ou camundongos, e está preparado dentro de uma capela de fluxo laminar em condições estéreis. Tempo total de coleta de tecido deve ser de 1 hr.
- Tome geraram filhotes saudáveis e bem nutridos (tamanho da leitegada ~ 10; presença de um 'spot de leite abdominal) em 1-2 dias pós-natal (PND). Segure um filhote de cachorro gentilmente pelo focinho, deixe-o solto, e rapidamente decapitar na base do pescoço com uma tesoura afiada.
- Para a remoção do cérebro, remover a pele (dois laterais cortes de tesoura), cortar o crânio aberto com tesoura olho fino (1 sagital corte da linha média; um corte coronal no córtex / junção cerebelo). Back flip todos os 4 flaps crânio. Com uma espátula afiada, corte frontal através do bulbo olfatório, espátula antecedência caudalmente debaixo do cérebro. Levante cuidadosamente o cérebro fora do crânio e deixá-lo deslizar para dentro estéril, refrigerado, Gey solução para resfriamento rápido e armazenamento temporário. Repita o 3.2 Passos para 3,3 para 2 cérebros mais (tempo total: <20 min).
- Para obter VTAtecidos, transferência de cérebros em uma placa de Petri estéril, seca com uma espátula pequena. Continuar a suprimir o excesso de fluido, deslizando cada cérebro cerca de 1 cm de lado. Remover tronco cerebral por um corte, coronal vertical ao nível do cerebelo usando uma lâmina de barbear.
- Bloco de agar Glue para a montagem do disco para a estabilização mecânica do cérebro durante o corte procedimento. Coloque uma linha fina de superglue alguns milímetros à frente do bloco de ágar no disco (evitar tocar a cola agar).
- Usando uma pequena espátula, transferir e montar cada cérebro, pólo frontal para baixo. Assegurar que os pólos frontais são colados ao disco de montagem e que os lados ventral tocar o agar sem resíduos de cola, a fim de alcançar a estabilização mecânica adequada durante o corte e fácil levantar-off de fatias de corte.
- Cuidadosamente submergir e disco montagem segura com a montagem do cérebro em uma bandeja vibratome (por exemplo, Leica VT1000) cheias de estéril, solução Gey de refrigerados. Com uma lâmina de barbear cuidadosamente limpos (90% EtOH), cortar fatias coronal do mesencéfalo na freqüência mais alta vibração e relativamente baixa velocidade de avanço com uma espessura de 400 500 mm. Usando uma pipeta Pasteur invertido com bomba de sucção, transferência e recolher fatias contendo o VTA em 35 x 10 milímetros pratos cheios de Petri estéril, solução Gey de refrigerados (Fig. 1C; ver placa também coronal 18-20 em E22 em 15).
- Para seções córtex, repita os passos de 3,2-3,6, mas aplicar corte vertical entre o córtex eo cerebelo, e montar prosencéfalo com pole frontal para cima. Cerca de 3 cortes coronais (350 espessura mm) começando no nível do corpo estriado são coletados para dissecção córtex futuro.
- Usando uma faca feita de micro-lâminas de barbear quebrado, dissecar ~ 2 mm de largura seção coronal do córtex frontal e áreas do mesencéfalo contendo o VTA (Fig. 1C), sob um estereomicroscópio. Coletar secções de tecido separadamente em pratos pequenos (por exemplo, slides câmara) preenchidas com solução de Gey refrigerados.
4. Montagem Cortex e fatias de tecido VTA no MEA (Time: <1 h)
- Posição MEA em temperatura ambiente sob um microscópio estéreo com o arranjo de eletrodos em foco. Centro de uma gota de 25 mL de plasma no limpo, livre de poeira, e matricial estéril eletrodo. Usando espátulas pequenas, deslize cuidadosamente um córtex e seção VTA na gota de plasma.
- MEA lugar na placa de refrigeração, reorientar vista, deixe esfriar para ~ 15 s, em seguida, adicione 25 mL da trombina para a gota de plasma. Usando a trombina ponta da pipeta, cuidadosamente espalhar a mistura de plasma / trombina com pequenos movimentos circulares por todo o MEA. Não toque no conjunto de eletrodos diretamente quebradiços. Suavemente a posição do córtex sobre a matriz com margem dorsal ao longo da fileira segundo eletrodo da matriz. Desta forma, as camadas superficiais desenvolvimento acabará por cobrir o lembrete da matriz. O VTA é colocado adjacente à fronteira ventral da seção córtex (Figura 1D).
- Cap e vagamente fechar a câmara de MEA para reter umidade elevada, enquanto a assembléia MEA / cultura fica por ~ 5 min dentro do capô à temperatura ambiente para permitir plasma / trombina coagulação. Enquanto isso, repita o passo 4,1-4,3 por 3 culturas mais.
- Juntar cuidadosamente 600 mL de meio de cultura em pequenas gotas para a câmara de cultura usando uma seringa de 1 cc com 25 x 08/05 agulha.
- Firmemente fechar a câmara de MEA e coloque MEA / montagem cultura na bandeja de armazenamento de balanço dentro da incubadora (Fig. 1B). Para acelerar o procedimento, 3-4 MEAs pode ser montado em seqüências que se sobrepõem. Tempo de montagem por 12 MEAs deve ser <1hr.
- Depois de dois dias in vitro (DIV), adicione 10 ml de inibidor de mitose. Atualizar cultura da mídia em 60% em 4 DIV ea cada 4 dias depois.
5. Gravação eletrofisiológicos e Geração de estímulo
- Para estabelecer a relação entre deflexões significativas no potencial de campo local (LFP) ea tendência de neurônios para disparar potenciais de ação, depois de gravar a atividade ~ 1 semana espontânea 5,6 a 24 kHz para ~ 10 minutos de cada eletrodo da MEA (Hardware : MEA1060 w / circuito de corte, o ganho x1200, 12 bit A / D, faixa de 0-4096 mV, sistemas multicanais; Software: MC_Rack). Terra é fornecido através do eletrodo de aterramento interno ou externamente, adicionando uma célula Ag / AgCl-metade.
- Separar os LFP com um filtro passa-banda de 1 - 200 Hz pico da atividade extracelular (passa-banda 300 - 3000 Hz). Atividade de pico pode ser classificada em atividade única e multi-unidade, utilizando classificadores off-line pico (por exemplo, Plexon Inc.). Calcule pico provocou médias para cada eletrodo. Para as culturas do córtex, a maioria médias irá identificar desvios negativos LFP (nLFP) como o momento preferido de spiking neuronal na cultura.
- Calcular para cada eletrodo um limiar de -3 desvios-padrão do ruído (SD) a partir de vestígios LFP (Fig. 2), Determinar os horários de pico e amplitudes de nLFPs que cruzam limiar (Fig. 2B, C). Escolheu um tempo Dt bin (por exemplo, entre 2-8 ms) e identificar surtos nLFP espaço-temporal da matriz concatenando nLFPs de todos os eletrodos que estão no mesmo tempo combina sucessivas de Dt comprimento (Fig. 2D; para mais detalhes veja 2,4 , 5).
- A fim de identificar avalanches neuronal, calcular o tamanho de cada cluster nLFP, por exemplo, número de eletrodos ativos ou soma de nLFP amplitudes, construir um histograma tamanho e parcela em duas vezes logarítmica coordenadas. Para avalanches neuronal, a distribuição de tamanho segue uma lei de potência aproximada por uma linha reta em coordenadas logarítmicas double-2 (Fig. 2E, F). Ver 16 para os testes estatísticos sobre as leis de potência.
- Eliciar respostas evocadas no tecido, selecionando um eletrodo através do qual a corrente controlada estímulos com amplitude S são aplicados (gerador de estímulo STG 1008, Sistemas Multicanal). Para reduzir os danos eletrodo, use um alcance limitado, carga neutra estimulação de choques único com bipolar onda quadrada: 50 mS com amplitude-S seguido de 100 mS com amplitude + S / 2 e S entre 1-20 mA. Consulte o manual do proprietário para obter mais detalhes.
- Para gravar a faixa dinâmica, 9 de estímulo recorde registrado respostas LFP em 4 kHz taxa de amostragem em todos os eletrodos seguintes 500 ms após a estimulação. Use blanking circuito (Multi-channel de sistemas) que desliga os amplificadores de palco cabeça durante a estimulação para reduzir artefatos de estímulo e evitar a saturação pré-amplificador.
6. Resultados representativos:
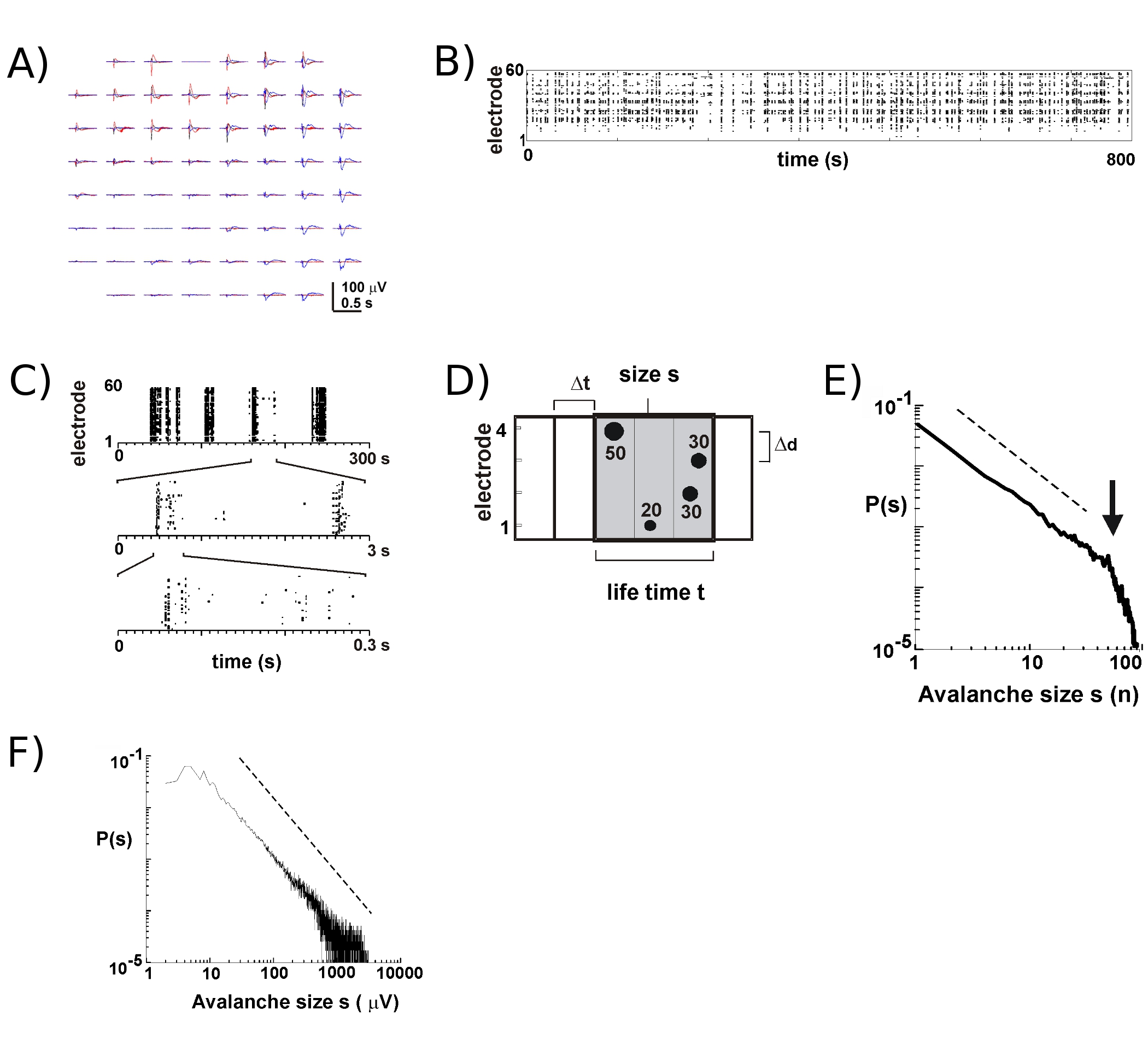
Com MEAs novo aproximadamente 8 - 9 de 10 culturas irão sobreviver por muitas semanas. A maioria dos nossos longo prazo gravações ocorrem no interior da incubadora no meio de cultura, o que nos permite acompanhar o desenvolvimento das culturas individuais ao longo de muitas semanas 5. Com base em nossas experiências, as gravações podem ser confiavelmente LFP obtidos com MEAs usado por mais de 100 dias da cultura. Em contraste, a atividade pico extracelular é mais confiável medida com MEAs relativamente novos (<40 dias de cultura). Em um experimento típico, nós transferimos uma MEA da bandeja de armazenamento (Figura 1B, direita) para a bandeja com o estágio de cabeça em anexo (Figura 1B, à esquerda) mantendo a câmara de cultura selado. Para o córtex 5, córtex-VTA co-culturas 6, bem como no rato anestesiado 6 eo macaco acordado in vivo 7, disparo neuronal durante avalanches neuronal nas camadas superficiais ocorre predominantemente perto da deflexão pico negativo da LFP (nLFP ). Assim, a organização espaço-temporal de grupos neuronais sincronizados localmente pode ser estimada medindo-se a ocorrência de nLFPs no espaço e no tempo nos 17 matriz.
Atividade no MEA tende a emergir em clusters temporal, tal atividade que, em um eletrodo é acompanhada por atividade em outros locais. Formas de onda típico da LFP durante os períodos de tal atividade são mostrados na Figura 2A, por mais de plotagem 3 clusters ocorrendo vários segundos de intervalo. Para cada cluster, deflexões campo negativo pode ser visto em vários eletrodos dentro de uma janela de 1 s. Ao extrair picos nLFP que cruzam um limite de múltiplos SD negativo, a atividade na forma de nLFP horário de pico está convenientemente visualizada em um raster em que "colunas" de pontos representam perto nLFPs coincidentes em vários eletrodos (Fig. 2B). A organização espaço-temporal desta actividade é bastante complexo; "colunas", que aparecem mais ou menos homogêneo em baixa resolução temporal, são compostas de grupos separados com maior resolução temporal e assim por diante (Fig. 2C). Na verdade, a emergência de clusters nLFP espaço-temporal é altamente organizados em redes cortical. Mais especificamente, a organização é escala-invariante de avalanches neuronal. Isto é demonstrado através do cálculo da probabilidade de tamanhos de cluster em um Dt determinada resolução temporal. Aqui, clusters são compostas de nLFPs que ocorrem nas caixas mesmo tempo ou sucessivos (Fig. 2D). Quando o tamanho de tal um cluster é expressa em número total de nLFPs por cluster, ou nLFP amplitudes integrada por cluster, as distribuições de tamanho do cluster revela uma lei de potência, cuja inclinação foi mostrado para ser 2,4,5,7 -1,5 ( fig. 2E, F). Note-se que esta distribuição identifica uma ordenação escala-invariante de tamanhos de cluster que é a relação de tamanhos para kxs s, onde k é um fator constante, é k -1,5, que é independente de s. Esta organização lei de potência é independente do tamanho da matriz 2, a resolução temporal Dt 2, eo limiar usado para identificar desvios significativos nLFP 7. Como as escalas amplitude nLFP com o tamanho do grupo neuronal 7, a organização em escala invariante de nLFPs reflete uma escala-invariante, ou seja, fractal, o ordenamentof localmente sincronizado grupos neuronais que incluem todos os tamanhos.

Figura 1. (A) e superior Side pontos de vista do MEA com anel de vidro com rosca montado, e tampa correspondente. (B) Vista do interior da incubadora. Esquerda: headstage montagem permitindo a gravação de uma única cultura em incubadora. Direita: Bandeja segurando MEAs numerosos para o crescimento da cultura. Rodas laterais: pisar dispositivo motor controlado de balanço para alternando fase submersa e-atmosfera expostos necessários para o crescimento da cultura. (C) Desenho esquemático para as fatias de rato usado para coronal córtex-VTA co-culturas. Seções Cortex (esquerda) e seções mesencéfalo (médio, direito), contendo a área tegmental ventral VTA (VTA; cinza) são obtidos através do corte ao longo das linhas quebradas. Ctx: córtex; ui: substância branca; cpu: striatum; VTA: pons: área pontina. Veja também correspondente placas coronal 8, 18, e 20 por 15. (D) Colocação e crescimento de uma única córtex-VTA co-culturas no MEA e seu desenvolvimento ao longo dos nove primeiros DIV cultura. Observe o achatamento da cultura e da sua expansão progressiva da matriz. Peças de tecido refletivo indicam células degeneradas e restos de tecido. Tecido saudável é opaco e acinzentado num transiluminador com luz visível.

Figura 2. Avalanches Neuronal na cortical culturas organotípicas. (A) Overplot de três período de atividade espontânea na matriz, separados por alguns segundos. Note-se que cada período de atividade consiste em deflexões LFP negativo sobre vários eletrodos no array (cada cor rótulos um período de atividade). (B) os horários de pico negativo de nLFPs de cada eletrodo são montados em um raster de atividade. "Column' estruturas semelhantes indicado períodos de atividade síncrona próximo. (C) Note que as colunas que parecem altamente sincronizados em uma escala de tempo consistem de várias colunas nas escalas maior temporal (3 escalas temporais mostrado). (D) representação esquemática do algoritmo avalanche neuronal. Em um tempo de 2 2 x eletrodo pico variedade e amplitude de desvios negativos LFP (nLFP) cruzar um limite de x-SD do ruído são identificados. Organização espaço-temporal de nLFPs é agrupada em caixas de tempo sucessivamente ativa de Dt largura. O tamanho de um cluster é identificado por tanto o número de sítios ativos, ou seja, com eletrodos nLFP (s = 4), ou a soma integrada de nLFP amplitudes (s = 130 mV). O tempo de vida é medida em múltiplos de Dt. (E, F) a lei de energia na distribuição de tamanho de cluster identifica clusters como avalanches neuronal. Note-se que a escolha de um determinado distâncias intereletrodos Δd para a matriz (aqui 200 mm) introduz uma Dt específico em que a dinâmica deve ser observada. Mais especificamente, a relação em que Δd / Dt se aproxima da velocidade de propagação média na rede, em que a inclinação α da lei de potência é aproxima -1,5 por avalanches neuronal 2,4,5. Por favor, clique aqui para ver uma versão ampliada da figura 2.
Discussion
1. Questões técnicas:
- Técnica estéril. A preparação dos MEAs e preparação cultura são todos realizados dentro de uma capela de fluxo laminar em condições estéreis. Antibióticos, que afectam a actividade neuronal, não são usados a qualquer momento durante o processo de preparação e cultura.
- Plasma / trombina de coagulação e adesão do tecido sobre a sobrevivência MEA. Tissue na MEA exige um cuidadoso equilíbrio entre o tempo necessário para plasma / trombina de coagulação e do tempo de exposição do tecido para a atmosfera. Um tempo de coagulação curto riscos de descolamento prematuro de seções da MEA, enquanto que a exposição prolongada a atmosfera provoca a degeneração dos tecidos. Porque a força da solução de trombina determina a velocidade do processo de coagulação, é um parâmetro muito importante para o sucesso anexando, culturas saudáveis para a superfície MEA. Obtemos os melhores resultados usando 1.000 unidades (1KU; 1 unidade NIH = 0,324 ± 0,073 mg). Importante, mistura incompleta dos resultados plasma / trombina na coagulação solução espacialmente heterogêneo promovendo plasma rupturas durante o cultivo. "Buracos" Estes plasma afetar gravemente a saúde, cultura e destacar a cultura de microeletrodos, comprometendo a qualidade da gravação eletrofisiológico. Trabalhando com placas de resfriamento durante o MEA / tecido montagem retarda a coagulação e permite a mistura homogênea da solução de plasma / trombina e bom posicionamento das seções do tecido.
Da mesma forma, pela adição de meio de gotículas única para a câmara de cultura para a cultura submergir depois de apenas 5 minutos de coagulação, reduz muito o risco de descolamento do tecido devido a tensões superficiais. Uma cultura de sucesso vai achatar a MEA e ligeiramente expandir mostrando crescimento saudável durante semanas, sem quaisquer sinais principais de contacto com o tecido-MEA incompleta ou degeneração do tecido (por exemplo, Fig. 1D).. - Facas dissecção dos tecidos. Micro melhoraram muito o nosso processo de dissecção. Usamos lâminas de duplo fio de navalha (Ferramentas Ciência Fine - quebrável lâminas de bisturi - 100.050-00), da qual nos separamos ~ 2 mm de largura 'blades' usando um alicate, e mantê-los com um suporte de bisturi. Amostras de tecido dissecado desde a corte coronal usando um movimento liso, firme vertical da lâmina, assim, reduzindo o estresse mecânico devido a puxar o tecido melhorar a saúde geral da cultura.
- Resfriamento tecidual. Resfriamento adequado das fatias e cortes de tecidos durante a preparação é essencial para o sucesso da cultura. Usamos feito por placas frias construído de elementos Peltier anexado por baixo para um disco de metal. O calor produzido pelo elemento Peltier é removida através de perfusão de água fria. Isso reduz muito o tempo de preparação e padroniza refrigeração durante cada estágio da preparação (Dold Labs e Engenharia 131 Plantation Dr. Seguin, TX 78155;. (830) 560-1471)).
- Incubadora condição. A incubadora custom-made, com condições precisas de balanço interno tem sido crucial para o nosso sucesso no cultivo de fatias em MEAs. Baseado em um original em casa design by Multichannelsystems (atualmente não disponível comercialmente), o dispositivo de balanço interno consiste em bandejas que estão ligados a duas rodas laterais de grandes dimensões. Motores passo a passo e controles computadorizados permitem uma trajetória de balanço preciso (ângulo de balanço, balançando, velocidade e pausas intermitentes). Em última análise, as culturas fatia precisam ser expostos à atmosfera e meio de cultura em alternância lento. A abordagem tradicional é colocar culturas fatia em tubos estreitos que girar lentamente por ali maior eixo. Aqui, rotação lenta não produz estresse mecânico devido à rotação em si, ea velocidade de rotação é alta o suficiente para obter um ótimo 'alimentação / respiração "ciclo de cerca de 5-10 minutos de duração. O interior mais compacta da câmara de MEA, seu volume total de ~ cc 2, eo volume pequeno médio necessário para a cultura condicionado média pelo tecido em si, representa um desafio significativo. Ao balançar o MEA entre ± ~ 70 ° de ângulo (tempo de ciclo: ~ 200 s), diminuindo a velocidade de balanço como as transições cultura entre a fase líquida ea atmosfera, e travar o balanço nos ângulos extremos de exposição prolongada à atmosfera tem sido essencial para a sobrevivência da cultura.
2. Idade de desenvolvimento de culturas para estudar avalanches córtex neuronal
Aguda fatias de córtex de ratos são comumente tomadas no PND 0-1 e cultivadas por muitas semanas no MEA. Os primeiros estudos demonstraram claramente que as culturas única fatia córtex, após várias semanas in vitro, manter uma estrutura em camadas com tipos celulares identificáveis que podem ser facilmente comparado ao em classes de células vivo 18,18-21. A organização em camadas neste sistema in vitro tem sido usado para estudar convenientemente talâmicainervação do córtex durante o desenvolvimento 22-24, bem como para a condução de estruturas subcorticais, como o striatum 25,26. Na verdade, a especificidade na formação de conexões neuronais dentro e entre regiões do cérebro permite a construção de complexos sistemas in vitro que recapturar numerosos sistemas de projeção detalhada, por exemplo, as dos gânglios basais córtex-circuito 27-30.
Depois de 4-6 semanas em vitro, fatias única córtex 31 e fatias de córtex co-cultivados com striatum 26 ou 32 tálamo mostrar espontânea cima e para baixo estados tipicamente encontrados in vivo em ratos anestesiados uretano 33. A organização temporal da multa nesses estados-up traz a marca de nested-θ e γ-oscilações indicativo de uma rede eletrofisiologicamente maduro de neurônios piramidais e fast-spiking interneurônios GABAérgicos 31. Importante, na ausência de estimulação dos receptores D2 de dopamina, a maturação de parvalbumina positivo interneurônios corticais é adiada por cerca de 2 semanas em culturas fatia córtex 34. Em linha com esses achados, o tempo de desenvolvimento de nested-θ, β e γ-oscilações corresponde ao in vivo quando fatias de córtex são co-cultivados com a área tegmental ventral (VTA), que contém neurônios dopaminérgicos projetam para o córtex 6.
Esses estudos indicam que quando se estuda avalanches neuronal, que dependem fortemente da inibição GABAérgica rápido maduro e estão localizadas nas camadas superficiais do córtex 2,4, muito cuidado deve ser tomado para assegurar a maturação adequada do tecido cortical. Enquanto avalanches neuronal em culturas surgem córtex única ao longo tempo de 2-5 semanas 4, ao exigir um curso de tempo de desenvolvimento que é comparado com o desenvolvimento in vivo, fatias de córtex precisa de uma estimulação adequada do receptor de dopamina, por exemplo, por fatias cultura co-cortex com o VTA 6.
Disclosures
Não há conflitos de interesse declarados.
Acknowledgments
Este estudo foi financiado pela Divisão de Programa de Pesquisa Intramural (dirp) do Instituto Nacional de Saúde Mental, National Institutes of Health.
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}