

Summary
皮質における臨界状態のダイナミクスを示す神経雪崩、すなわちスケール不変時空間アクティビティのバーストを、勉強するための確実な方法。雪崩は、正確に制御された条件下で平面統合マルチ電極アレイ(MEA)との活動の長期的な測定を可能にする培養皮質の浅層の開発に自発的に出現する。
Abstract
皮質はさらに、特定の入力またはモーターの出力がない状態で、自発的にアクティブになります。開発中は、この活動は、皮質の細胞型と神経の接続1の形成の遊走と分化に重要である。成熟した動物では、継続的な活動は、過去と感覚刺激がシームレスに将来の行動を計算するために統合された動物の現状を反映している。このように、継続的な、すなわち自発活動の組織を明確に理解することは、皮質の機能を理解することが前提条件です。
数々の記録技術は、皮質における継続的な活動が、その個々の活動一過性に細胞外微小電極で局所電場電位(LFP)で検出することができる大規模なイベントに合計、または脳波(EEG)、脳磁気図(MEGの多くのニューロンから構成されていることを明らかに)、そして機能的磁気共鳴画像(fMRI)からBOLD信号。メゾスコピックスケール(ニューロン数千)での高時間および空間分解能で神経集団の活動を研究するときにLFPは、現在の選択の方法です。細胞外微小電極では、局所的にマイクロボルトから数百までのLFPの急激なたわみの空間的neighboredニューロンの結果の活動を同期化。微小電極のアレイを使用する場合は、このような偏向の団体は、便利な空間と時間で監視することができます。
神経雪崩は脳2,3の継続的なニューロン活動のスケール不変時空間構成について説明します。 in vitroで 4,5 で 、麻酔ラット6のin vivoで 、と目を覚ましサル7に設立されたとして、それらは皮質の浅層に固有のものです。重要なことは、理論的実証的研究2,8-10両方の神経雪崩の情報伝達と情報の処理を最適化する皮質の絶妙バランスの臨界状態のダイナミクスを示すことを示唆している。
彼らは正確に制御された条件下で雪崩の活動の安定的な録音を可能と神経雪崩の開発、保守、および規制のメカニズムを研究するためには、in vitroでの製剤で 、非常に有益です。現在のプロトコルは、平面、統合された微小電極アレイ(;も11-14を参照してMEA)上に成長した器官皮質の培養、すなわちスライス培養で浅層の開発を活用することによって、in vitroで神経雪崩を勉強する方法について説明します。
Protocol
1。長期録画用MEAと滅菌、密閉式ガラスチャンバー
- 安全性とタイトな培養チャンバーの閉鎖に必要なテフロンプラスチックキャップ(エースガラス)、、とスレッド化ガラスシリンダー(Aceglass)スレッドの底部(図1A、B)から約2 mmにカットされています。水(3倍)と200プルーフエチルアルコールで5分間煮沸して洗浄して清浄なガラスリングは、乾燥させる。
- MEAの表面にガラスリングを取り付けるために必要な一定分量のシリコンソリューション。 Sylgard 184シリコーンエラストマーのキットの&Bは、徹底的に、-20℃で1mlのアリコートに店舗、気泡を除去するために15分間放置します部品の15 mlを混ぜて
- MEAに接着剤のガラスリング(8 × 8のグリッドのw /内部接地電極、ラット/マウス用の30μmの電極の直径、100分の200μmの電極間の距離)(図1A、B)。小さなゲージの針付き注射器でシリコンを1 ml(23℃)を取り上げる。 〜60℃のホットプレート上で2時間 - ガラスリング、MEA上の中央のガラスリング、強力なシール用リングの外側を、シリコンの層を追加適用の未研磨の切断面にシリコーンを適用、1に治すことができます。
- 紫外線にチャンバーとキャップ内部の10分露光に続いて、70%アルコールに続いて(最後のためにアルコールで10分間放置しますリンス3 ×)、脱イオン水でリンス3倍で層流フードでMEAチャンバーとチャンバーキャップを滅菌。オートクレーブMEA室(120℃、ウェット、45分)と乾燥させる。
- ポリ- D -リジンでコーティングMEAの表面培養チャンバー内の。用ではなく、親油性である新しい多国間環境協定、、電極のグリッドからのソリューションの繰り返し滴の吸引コート。使用される多国間環境協定の場合は、ソリューションを持つチャンバーの底部をカバーする、余分な液を吸引し、層流フード内無菌条件下で蒸発させる。ストレージおよび将来使用するためにシールMEA室にキャップを取り付けます。
2。器官培養の調製と成長に必要な成分
- 滅菌ペトリ皿(ファルコン100 X15;〜5mmのレベル)に注ぐ、0.9%NaCl中で無菌の寒天を溶かし、冷ます、およびストレージ用のパラフィルムで滅菌ラップ。用固形寒天から20 × 10 × 5mmのブロックをカット。
- パッケージングと店舗スーパー接着剤、例えば、DevConのスーパー接着剤IIは、無菌性を維持するために層流フードの内部で、開く前に、70%エタノールでダウン拭った。
- 脱イオン化文化の40mlの水(Sigma社)にグルコース40gを添加することにより50%D -グルコース(SIGMA超、G7528)を準備する。 -20℃で2 mlのアリコートの店
- Geyの平衡塩類溶液500mlに50%D -グルコースの4 mlを加え、そして使用前に冷凍庫でスラッシュ(液体/氷結晶の混合物)に冷やします。
- 5ミリリットルの脱イオン文化の水の再構成の鶏の血漿(優しく振る、気泡の形成を避けるため)、ソリューションが5に甘んじることができます - 10分、静かに渦巻き、そして滅菌シャーレに明確なコンテンツをdecantate。滅菌フィルター(0.22μm孔フィルター、蛋白質)-20℃でプラズマのソリューション、クリオチューブに分注し350μlの(NuncTM)、店舗
- クリオチューブ(NuncTM)に応じて牛血漿からの再構成のトロンビン、滅菌フィルター(0.22μm孔フィルター)、アリコート40μlの、-20℃で保存します。ソリューションを操作するための、Geyの平衡塩類溶液W / D -グルコースの375μlのトロンビン溶液の40μlを希釈する。
- ウマ血清、200ミリリットル基礎培地イーグル、50%グルコースと200 mMのL -グルタミンを2ml 4mlのが追加される100ミリリットルハンクス緩衝生理食塩溶液100 mlを混合して培養液400 mlを調製。 4℃で8週間100ミリリットルパイレックスのボトル℃と - 4を格納することができます
- 0.3mmのウリジン、0.3mMのARA - Cシトシン-β- D -アラビノフラノシド、および0.3mMの5 - フルオロ-2' - デオキシウリジン、滅菌フィルタ、分注して200μlの、6、-20℃で保存を混合することによって有糸分裂阻害剤を準備する - 12ヶ月。
3。皮質と腹側被蓋野(VTA)組織の解剖(時間:<1時間)
- プロシージャ利回りラットやマウスから〜12共培養のための皮質とVTAの組織切片、および無菌条件下で層流フードの内部に用意されています。組織採取のための合計時間は1時間以上でなければなりません。
- 2生後日数(PND) - 1で、健康的な、よく栄養仔(腹部"ミルクのスポット"の存在仔数〜10)を取る。鼻でそっと子犬を保持し、それが自由にハングアップすることができます、とすぐに切れるはさみで首の基部に首を切る。
- 脳を除去するため、(2つの側面シザーカット)皮膚を取り除く、細かい目のはさみ(1矢状正中線のカット、皮質/小脳ジャンクションで1冠状カット)でオープン頭蓋骨をカット。すべての4頭蓋骨のフラップをバックフリップ。シャープスパチュラで、嗅球、尾側の脳の下に事前にへらを通して正面を切る。ゆっくりと頭蓋骨から脳を持ち上げ、それが急速な冷却と一時記憶用の滅菌、チルド、Geyの溶液中にスライドさせてみましょう。さらに2頭(:<20分合計時間)のためのステップ3.2から3.3を繰り返します。
- VTAを取得するには組織は、小さなヘラを使って滅菌、乾燥ペトリ皿の上に脳を転送する。さらに静かに横に約1cmの各脳をスライドさせて余分な水分を取り除く。カミソリの刃を使用して小脳のレベルで冠状、垂直カットで脳幹を削除します。
- 手順をスライス中に脳の機械的な安定化のためにディスクをマウントする場合、接着剤の寒天ブロック。瞬間接着剤の細い線を配置し、ディスク上の寒天ブロックの前に数ミリメートル(寒天に触れない接着剤を避けるため)。
- 小さなヘラを使用して、ダウン、前頭極を転送し、各脳をマウントします。前頭極が取り付けディスクに接着されていると腹側の側面は、中にカットスライスのカットと簡単なリフトオフ。適切な機械的安定を達成するために任意の接着剤の残留物なしで寒天に触れていることを確認してください
- 滅菌、冷却したGeyの溶液で満たされたビブラトームトレイ(例えばライカVT1000)で脳のアセンブリと慎重に沈めると、安全なディスクをマウント。慎重に洗浄かみそりの刃(90%エタノール)で、最も高い振動周波数での中脳の冠状スライスをカットし、400 500μmの厚さで、比較的低い前進速度。吸引器と反転パスツールピペットを用いて、(; - 15のE22で20にも冠状板18を参照して図1C)、滅 菌、冷却したGeyの溶液で満たされた35 x 10 mmのペトリ皿にVTAを含むスライスを転送して収集する。
- 皮質のセクションは、手順3.2の - 3.6が、前頭極、最大で縦皮質と小脳の間のカット、およびマウントforebrainsを適用します。線条体のレベルから始めて約3冠状スライス(350μmの厚さ)は、将来の皮質の解剖のために収集されます。
- 壊れたかみそりの刃で作られたマイクロナイフを使用して、実体顕微鏡下でVTAを(図1C)を含む前頭皮質と中脳領域の約2 mm幅の冠状断面を詳細に分析します。チルドGeyの溶液で満たされた小さな料理(例えばチャンバースライド)で別々に組織切片を収集する。
4。 MEA上のCortexとVTAの組織切片をマウントする(時間:<1時間)
- 焦点の電極アレイ付きステレオ顕微鏡下で、室温でMEAを置きます。中央クリーン、無塵、無菌電極アレイのマトリックス上にプラズマの25μlの液滴。小さなspatulaeを使用して、慎重にプラズマ液滴に皮質やVTAセクションをスライドさせます。
- 〜15秒のために寒さを聞かせ、表示する再び焦点、冷却板のMEAを置き、プラズマ液滴にトロンビン25μlを加える。トロンビンピペットチップを使用して、慎重にMEA間の小さな円形の動きとプラズマ/トロンビン混合物を広げる。直接脆性電極アレイを触れないでください。優しく配列の第2の電極の行に沿って背側縁を持つ配列に皮質を置きます。このように、発展途上浅層は最終的には配列のリマインダをカバーします。 VTAは皮質部の腹側の境界線(図1D)に隣接して配置されます。
- キャップと緩くMEA /文化アセンブリは、プラズマ/トロンビン凝固を可能にするために、室温でフードの内側〜5分間座っている間、高湿度を維持するためにMEAのチャンバーを閉じます。さらに3文化4.3 - 一方、ステップ4.1を繰り返します。
- 慎重に25 × 5 / 8針で1 ccシリンジを用いて培養チャンバーに小さな液滴内の培養液600μlを加える。
- しっかりとMEA室と場所MEA /インキュベーター(図1B)の内部ロッキングストレージトレイの文化のアセンブリを閉じます。に、3の手順をスピードアップ - 4のMEAは、重複する配列で組み立てることができます。 12多国間環境協定のための組立時間は<1時間でなければなりません。
- in vitroでの 2日間(DIV)の後に、有糸分裂阻害剤10μlを加える。 4 DIVおよびその後4日毎でのリフレッシュの培地60%。
5。電気生理学的記録と刺激生成
- MEAの各電極から〜10分の場合は24 kHzで約1週間5,6レコードが自発的な活動の後、ローカルフィールドポテンシャル(LFP)と活動電位を発生させるには神経細胞の傾向に大きなたわみとの関係を確立するには(ハードウェア:MEA1060 W /ブランキング回路、X1200ゲイン、12ビットA / D、範囲0から4096 mVの、マルチチャンネルシステム、ソフトウェア:MC_Rack)。グランドは内部接地電極を介して、または外部Ag / AgCl電極、半電池を追加して、提供されています。
- 細胞外スパイク活動から200 Hzの(バンドパス300 - - 3,000 Hz)の1のバンドパスフィルタでLFPを区切ります。スパイク活動は、さらにオフラインでスパイクの選別機(例えばPlexon社)を用いて、単一およびマルチユニット活動に分類することができます。スパイクは、各電極の平均値をトリガー計算する。皮質培養の場合は、最も平均値は、培養中のニューロンのスパイクの希望する時間として負のLFPのたわみ(nLFP)を識別します。
- (図2)それぞれの電極にLFPのトレースからのノイズ(SD)の-3標準偏差の閾値を計算する、クロスしきい値(図2B、C)そのnLFPsのピーク時と振幅を決定する。時間ビンΔtの(2の間など- 8ミリ秒)を選択し、長さΔt(図2Dの連続したタイムbinesの同じ内にあるすべての電極からnLFPsを連結することによって、アレイ上の時空間nLFPクラスタを識別し、詳細については、2,4を参照してください、5)。
- 神経雪崩を識別するためには、各nLFPのクラスタのサイズを計算する、アクティブ電極またはnLFP振幅の合計の例数は、サイズのヒストグラムを構築し、二重対数座標にプロット。神経雪崩の場合は、サイズ分布は、二重対数座標2(図2E、F)の直線で近似べき乗則に従う。電源の法律上の統計的検定のための16を参照してください。
- 振幅Sと電流制御の刺激が(刺激発生器STG 1008、マルチチャンネルシステム)適用されるときに通過する電極を選択することにより、組織における誘発反応を引き出す。 - 200μA振幅は- Sで50μsの10の間の振幅+ S / 2とSで100μsの間に続く:電極の損傷を減らすために、限られた範囲、バイポーラ方形波形を持つ単一の衝撃の電荷中性の刺激を使用してください。詳細は取扱説明書を参照してください。
- ダイナミックレンジ9録音するには、記録的な刺激は、刺激後500ミリ秒を、以下の全ての電極上に4 kHzのサンプリングレートでLFPの応答を記録した。刺激アーチファクトを低減し、プリアンプの飽和を防止するために刺激時のヘッドの段増幅器を切断するブランキング回路(マルチチャンネルシステム)を使用してください。
6。代表的な結果:
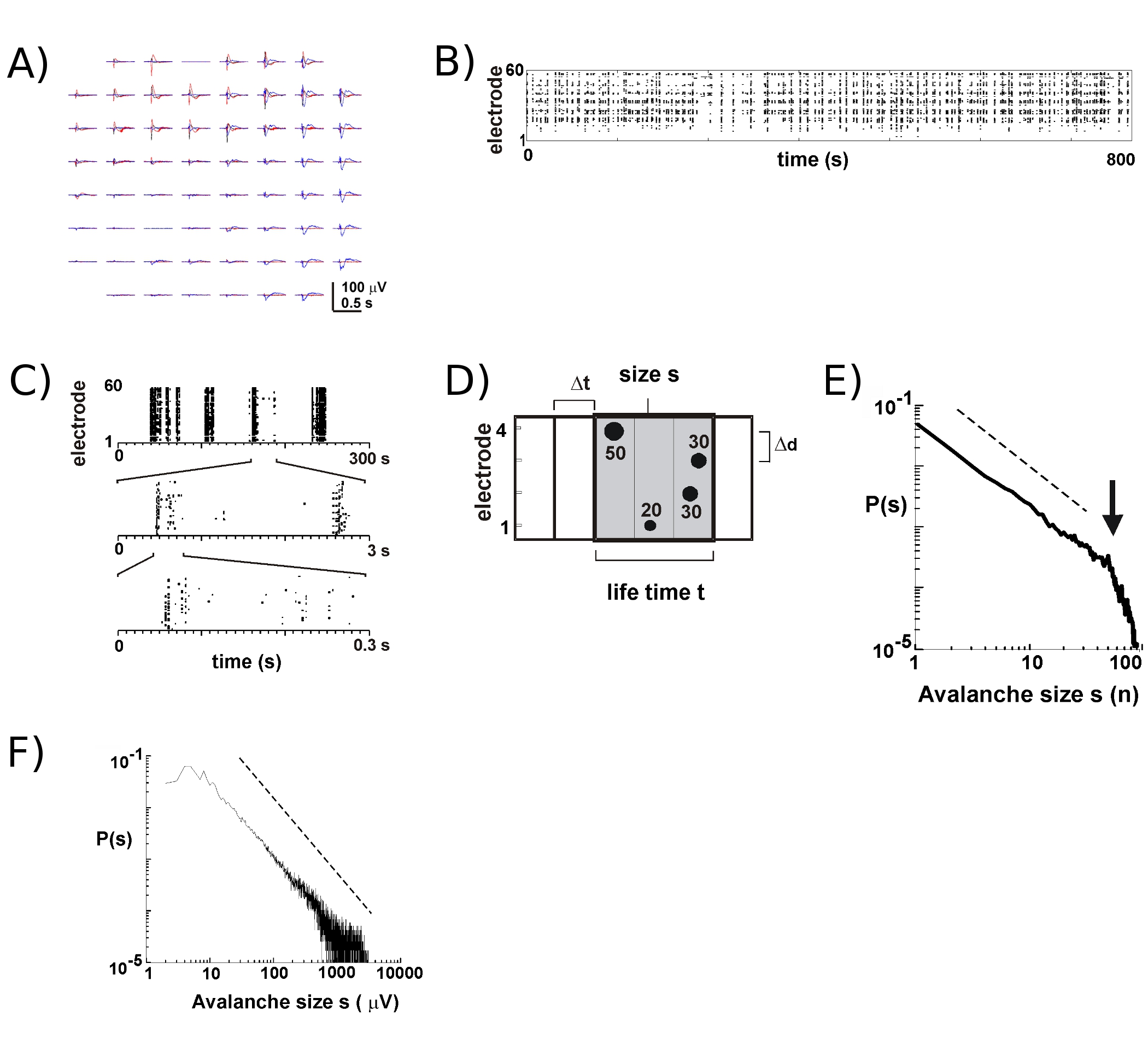
約8新しい多国間環境協定との - 9 10文化の何週間も存続します。私たちの長期的な録音のほとんどは、私たちは何週間のコース5を介して個々の文化の発展をフォローできるようにする培地で培養器の内部で行われる。我々の実験に基づいて、LFPの記録は確実に100以上の文化の日のために使用される多国間環境協定で得ることができる。対照的に、細胞外スパイク活動をより確実に、比較的新しい多国間環境協定(<40文化の日)で測定されます。典型的な実験では、我々は、培養容器は、密閉式保つ付属のヘッドステージ(図1B、左)とトレイに収納トレイ(図1B、右)からMEAを転送する。皮質5の、皮質- VTA共培養6、同様に麻酔をかけたラット6、in vivoで 7 の目を覚まし猿、浅層の神経雪崩時の神経発火のように、主にLFPのピーク陰性波に近い発生します(nLFP )。従って、局所的に同期神経群の時空間の組織は、アレイ17上の空間と時間のnLFPsの発生を測定することによって推定することができます。
MEA上のアクティビティは、一時的なクラスタに出現する傾向がある、一方の電極ではそのようなその活動が他の部位の活動が付属しています。このような活動期間中のLFPの典型的な波形を離れて数秒を発生する3つのクラスタをプロット上で、図2Aに示されています。各クラスタの場合は、負のフィールドのたわみは、1秒のウィンドウ内でいくつかの電極で見ることができます。複数の負のSDの閾値を越えるnLFPピークを抽出する場合、nLFPピーク時の形での活動は、ドットの"列"は、様々な電極(図2B)で一致nLFPs近く表しているラスターに便利な可視化である。この活動の時間的空間的組織は、かなり複雑で、低時間分解能で多かれ少なかれ均一な表示"の列は"、、より高い時間分解能など(図2C)で別々のクラスタで構成されています。実際には、時空間nLFPクラスターの出現は非常に皮質ネットワークで構成されています。具体的には、組織は、神経雪崩のスケール不変です。これは、与えられた時間分解能Δtにおいてクラスタサイズの確率を計算することによって実証される。ここで、クラスタは、同じまたは連続した時間ビン(図2D)で発生するnLFPsで構成されています。このようなクラスタのサイズがクラスタごとにnLFPs、またはクラスタごとに統合されたnLFP振幅の合計数で表現されている場合、クラスタのサイズ分布は、その傾き-1.5 2,4,5,7であることが示されている電力の法則を、明らかに(図2E、F)。この分布はスケール不変kは定数因子であるKXS、にサイズsの比であるクラスタサイズの順序を識別する、秒から独立している-1.5 kは、であることに注意してくださいこのベキ法則の組織は、アレイ2の大きさ、時間分解能Δtの2、および重要なnLFPのたわみ7を識別するために使用するしきい値とは無関係です。神経グループサイズ7とnLFP振幅スケールは、nLFPsのスケール不変の組織はスケール不変、すなわちフラクタル、順序Oを反映しているので、fは局所的にすべてのサイズを含む神経細胞のグループを同期。

図1。マウントされているスレッドのガラスリングを持つMEA、および対応するキャップの()側と上面図。 (B)インキュベーターのビューの中。左:インキュベーターの条件の下で、単一の培養液からの録音を可能にするヘッドマウント。右:培養増殖のために数々のMEAを保持するトレイ。側の車輪:培養増殖に必要な水中と大気にさらされたフェーズを交互にするためのモータ制御のロッキングデバイスのステッピング。皮質- VTA共培養に用いる冠状ラットスライス用(C)模式図。 VTA腹側被蓋領域(VTA、灰色)を含む皮質のセクション(左)と中脳のセクション(中央、右)は、破線に沿って切断することによって得られる。 CTX:皮質、WM:白質、CPU:線条体、VTA:橋:橋エリア。 15日までに、対応する冠状板8、18、および20を参照してください。 (D)の配置とMEAと文化の中で最初の9 DIV上に、その開発上の単一の皮質- VTAの共培養の成長。文化と、アレイ上で、その進歩的な拡大の平坦化に注意してください。反射組織部分では、変性細胞と壊死組織片を示す。健康な組織は、可視光で透視下で不透明と灰色です。

図2。皮質器官培養において神経雪崩。 ()配列での自発活動の三期間の重ねてプロットは、数秒で区切られています。各アクティビティの期間は、アレイ上に多数の電極にマイナスのLFPのたわみ(各色は、つのアクティビティの期間をラベル)で構成されていることに注意してください。 (B)の各電極からnLFPsの負のピーク時間は、活動のラスターに組み立てられます。 "Column'状の構造は、近くの同期活動の期間を示した。 (C)高度に一度のスケールで同期される列は、より高い時間的スケール(3時間スケールが示すように、)で複数の列で構成されていることに注意してください。神経雪崩のアルゴリズムの(D)模式図。ノイズのSD - xのしきい値を超える負のLFPのたわみ(nLFP)の2 × 2電極アレイのピークの時間と振幅で識別されます。 nLFPsの時間的空間的組織は、幅Δtの連続的アクティブタイムビンにクラスタ化されている。クラスタのサイズがnLFPと活性部位の数、すなわち電極のどちらかによって識別される(S = 4)、またはnLFP振幅の統合された和(S = 130μV)。寿命時間は、Δtの倍数で測定されます。クラスタのサイズ分布の(E、F)べき乗則は、神経雪崩のようにクラスタを識別します。配列(ここでは200μm)のためにΔdを特定の電極間距離の選択は、ダイナミクスが観察されるべきで、特定のΔtを紹介することに注意してください。具体的には、ΔD/Δtはべき乗則の傾きαがあるときの、ネットワーク内の平均伝播速度を近似した比率は、神経雪崩2,4,5に対して-1.5近似している。してくださいここをクリックして図2の拡大バージョンを参照すること。
Discussion
1。技術的な問題:
- 無菌テクニック。多国間環境協定と文化の準備の準備はすべて無菌条件下で層流フード内で実行されます。神経活動に影響を与える抗生物質は、準備および文化プロセス中にいつでも使用されていません。
- MEA上のMEA。組織の生存上のプラズマ/トロンビン凝固や組織の付着は、プラズマ/トロンビン凝固及び大気への組織暴露の時に必要な時間の間で注意深くバランスを取る必要。大気中に長時間暴露は、組織の変性をトリガしながら短い凝固時間は、MEAからセクションの早期の剥離をリスク。トロンビン溶液の強さは、凝固プロセスの速度を決定するので、それは成功したMEAの表面に、健全な文化を取り付けるための非常に重要なパラメータです。我々は、1000単位(; 1 NIHのユニット= 0.324 ± 0.073 mgの1,000個ロット)を使用して最良の結果を得る。重要なことは、培養中にプラズマ破裂を促進する空間的に不均質凝固におけるプラズマ/トロンビン溶液の結果の不完全混合。これらのプラズマ"の穴は"深刻な文化の健康に影響を与えると微小電極から文化を切り離し、それによって電気生理学的記録の品質を損なう。 MEA /組織の組み立て時の冷却板での作業凝固を遅らせ、プラズマ/トロンビンソリューションおよび組織切片の適切な配置の均質な混合が可能になります。
同様に、凝固のわずか5分後に水没文化に培養チャンバーに、単一液滴の培地を添加することにより、大幅に表面張力に起因する組織の剥離の危険性を低減します。成功する文化がMEAに平らにし、わずかに不完全な組織- MEAの接触または組織の変性(例えば、図1D)のいずれかの主要な徴候なしに、数週間のための健全な成長を示す展開されます。 - 組織の解剖。マイクロナイフは非常に私たちの解剖のプロセスを改善している。我々は約2 mm幅の"ブレード"を使用してペンチを分割し、メスホルダーでそれらを保持しているから、我々は、ダブルエッジのカミソリの刃(100050〜00 - - 壊れやすいメス刃ファイン科学のツール)を使用してください。それによって大幅に起因する文化の健康を改善する総合的な組織を引っ張るために機械的ストレスを低減するブレードの滑らかな、縦の企業の動きを使用して冠状スライスから解剖組織サンプル。
- 組織の冷却。準備中にスライスし、組織切片の適切な冷却は、文化の成功に不可欠です。我々は、金属板の下側取り付けられたペルチェ素子で構成されたカスタムメイドのコールドプレートを使用してください。ペルチェ素子によって発生する熱は、冷水の血流を介して削除されます。これは、大幅に準備時間を短縮し、準備の各段階で冷却を標準化する(DOLD研究所エンジニアリング131プランテーション博士セガン、TX 78155;(830)560〜1471))。
- インキュベーターの条件。カスタムメイドのインキュベーター高精度の内部揺動条件では、MEAの上のスライスを育成における当社の成功のための最も重要な鍵だった。 (現在ではない市販)Multichannelsystemsの社内デザインのオリジナルに基づいて、内部のロッキングデバイスには二つの大きな側面の車輪に接続されているトレイで構成されています。ステッピングモータとコンピュータのコントロールは、(速度の揺動角度を揺らし、、と断続的に一時停止します)正確なロッキングの軌跡を可能にする。最終的に、スライス培養は遅い交替の雰囲気と文化メディアに公開する必要があります。伝統的なアプローチは、徐々にそこに長い軸に沿って回転する細いチューブにスライス培養を配置することです。ここで、ゆっくりと回転は回転自体に起因する機械的ストレスを生成しませんし、回転速度が約5の最適な"摂食/呼吸"サイクルを得るために十分に高いです - 10分の時間を。 MEA室、〜2 ccのその合計量、そして組織自体が培地空調に必要な小規模な培養培地の容積のよりコンパクトなインテリアは、重要な課題となっています。 ±〜70 °の角度(サイクルタイム:〜200秒)の間にMEAを揺らすことによって、している液相と大気との間の文化の遷移に応じ揺動速度を減速し、大気への長時間曝露のために極端な角度で揺動を停止文化の生存に必須であった。
2。神経雪崩を研究するために皮質の文化の発達年齢
ラットの大脳皮質からの急性スライスは一般的にPND 0で取られる - 1とMEA上で多くの週間培養する。初期の研究は明らかに単一の皮質の切片培養は、in vitroで数週間後、容易にin vivoでの細胞のクラス18,18-21 のに比較することができる識別可能な細胞の種類と階層構造を維持することを明らかにした。 in vitroの系でこの中の層状組織が 便利に視床を研究するために使用されています開発22〜24時の大脳皮質の神経支配、だけでなく、線条体25,26のような皮質下構造を駆動するための。実際には、脳の領域内および組織間の神経接続の形成の特異性は、例えば皮質-大脳基底核回路27から30までのもの、多数の詳細な投影システムを奪還in vitro系における複合体の構築が可能になります。
4の後に- in vitroでの6週間、単一の皮質スライス31および皮質スライスが線条体26または視床32と共培養自発的には、上下を示す典型的にはウレタン麻酔ラット33に生体内で使用されるステート 。これらのアップ状態の微細な時間的な組織では、錐体ニューロンと速いスパイクGABA作動性介在ニューロン31の電気生理学的に成熟したネットワークのネストされたθ-とγ-振動を示すの特徴を負いません。重要なのは、ドーパミンD2受容体刺激の非存在下で、パルブアルブミン陽性皮質介在ニューロンの成熟は、大脳皮質のスライス培養34に約2週間遅れで表示しています。皮質スライスをする投射ドーパミン作動性ニューロンを含む腹側被蓋野(VTA)、共培養しているときにこれらの知見に沿って、ネストされたθ-、β-およびγ-振動の発達時間のコースが生体内でそれに一致さ皮質6。
これらの研究は極めて成熟した高速なGABA作動性抑制に依存すると皮質2,4の浅層に配置されている神経雪崩を、勉強するときに、細心の注意が皮質組織の適切な成熟を 確保するために取られる必要があることを示している。神経雪崩が2の時の経過とともに、単一の皮質培養で発生しながら- 5週間4、in vivoでの開発のためにマッチした開発時間の経過を必要とするときに、皮質スライスは、共培養皮質スライスによって、例えば適切なドーパミン受容体の刺激を、必要VTA 6と。
Disclosures
利害の衝突は宣言されません。
Acknowledgments
この研究は、国立精神衛生研究所、国立衛生研究所の学内研究プログラムの部門(DIRP)によって賄われていた。
Materials
| Name | Company | Catalog Number | Comments |
| Integrated planar multielectrode array |  Multi Channel System MCS GmbH Multi Channel System MCS GmbH |
200/30iR-ITO-w/o | Titanium Nitrate (TiN) electrodes (30 mm diameter) have large surface resulting in low impedance ( ~1.5 kΩ at 1 kHz) and excellent wide-band recordings ( w/o -– without ring) |
| Chamber glass | www.aceglass.com | 7620-32 | Threaded glass cylinder |
| Chamber cap | www.aceglass.com | 7622-114 | Plastic cap with Teflon insert |
| Sylgard 184 | World Precision Instruments, Inc. | SYLG184 | Two-part silicone elastomer |
| Poly-D-lysine | Sigma-Aldrich | P6407-5mg | γ-irradiated, lyophilized powder, cell cultured tested. Reconstituted with 5 ml deionised water before use. |
| Gey’s Balanced Salt solution | Sigma-Aldrich | G9779-500mL | sterile filtered and cultured tested |
| chicken plasma | Sigma-Aldrich | P3266-5mL | Lyophilized, reconstitute with 5 ml deionized water before use. |
| thrombin | Sigma-Aldrich | T6634-1KU | from bovine plasma, lyophilized powder form. |
| horse serum | Sigma-Aldrich | H1138-100mL | donor herd, heat inactivated, cell culture tested |
| Basal Medium Eagle | Invitrogen | 21010-046 | 1x, 500 ml - (+) Earle’s Salts, (-) L-glutamine), |
| Hank’s Buffered Saline Solution | Invitrogen | 24020-117 | 500 ml - (+) Magnesium, (+) calcium, w/phenol red) |
| Chamber slides | Lab-Tek | 177429 | |
| Uridine | Sigma-Aldrich | U3003 | |
| ARA-C cytosine-β-D-arabinofuranoside | Sigma-Aldrich | C6645 | |
| 5-fluoro-2’-deoxyuridine | Sigma-Aldrich | F0503 |
References
- Spitzer, N. C. Electrical activity in early neuronal development. Nature. 444, 707-712 (2006).
- Beggs, J. M., Plenz, D. Neuronal avalanches in neocortical circuits. J. Neurosci. 23, 11167-11177 (2003).
- Beggs, J. M., Plenz, D. Neuronal avalanches are diverse and precise activity patterns that are stable for many hours in cortical slice cultures. J Neurosci. 24, 5216-5229 (2004).
- Stewart, C. V., Plenz, D. Inverted-U profile of dopamine-NMDA-mediated spontaneous avalanche recurrence in superficial layers of rat prefrontal cortex. J. Neurosci. 26, 8148-8159 (2006).
- Stewart, C. V., Plenz, D. Homeostasis of neuronal avalanches during postnatal cortex development in vitro. J. Neurosci. Meth. 169, 405-416 (2007).
- Gireesh, E. D., Plenz, D. Neuronal avalanches organize as nested theta- and beta/gamma-oscillations during development of cortical layer 2/3. Proc. Natl. Acad. Sci. U. S. A. 105, 7576-7581 (2008).
- Petermann, T. Spontaneous cortical activity in awake monkeys composed of neuronal avalanches. Proc. Natl. Acad. Sci. U. S. A. 106, 15921-15926 (2009).
- Kinouchi, O., Copelli, M. Optimal dynamical range of excitable networks at criticality. Nature Physics. 2, 348-351 (2006).
- Shew, W. L., Yang, H., Petermann, T., Roy, R., Plenz, D. Neuronal avalanches imply maximum dynamic range in cortical networks at criticality. J. Neurosci. 29, 15595-15600 (2009).
- Shew, W. L., Yang, H., Yu, S., Roy, R., Plenz, D. Information capacity is maximized in balanced cortical networks with neuronal avalanches. J Neurosci. 5, 55-63 (2011).
- Karpiak, V., Plenz, D. Preparation and maintenance of organotypic cultures for multi-electrode array recordings. Current Protocols in Neuroscience. 1, 6-6 (2002).
- Hammerle, H., Egert, U., Mohr, A., Nisch, W. Extracellular recording in neuronal networks with substrate integrated microelectrode arrays. Biosens. Bioelectron. 9, 691-696 (1994).
- Nisch, W., Bock, J., Egert, U., Hammerle, H., Mohr, A. A thin film microelectrode array for monitoring extracellular neuronal activity in vitro. Biosens. Bioelectron. 9, 737-741 (1994).
- Egert, U. A novel organotypic long-term culture of the rat hippocampus on substrate-integrated multielectrode arrays. Brain Res. Protoc. 2, 229-242 (1998).
- Altman, J., Bayer, S. A. Atlas of Prenatal Rat Brain Development. , CRC Press. Boca Raton. (1995).
- Clauset, A., Shalizi, C. R., Newman, M. E. J. Power-law distributions in empirical data. arXiv. , (2009).
- Plenz, D., Thiagarajan, T. C. The organizing principles of neuronal avalanches: cell assemblies in the cortex? Trends Neurosci. 30, 101-110 (2007).
- Cäser, M., Bonhoeffer, T., Bolz, J. Cellular organization and development of slice cultures from rat visual cortex. Exp. Brain Res. 477, 234-244 (1989).
- Cäser, M., Schüz, A. Maturation of neurons in neocortical slice cultures. A light and electron microscopic study on in situ and in vitro material. J. Hirnforsch. 33, 429-443 (1992).
- Götz, M., Bolz, J. Development of vasoactive intestinal polypeptide (VIP)-containing neurons in organotypic slice cultures from rat visual cortex. Neurosci. Lett. 107, 6-11 (1989).
- Götz, M., Bolz, J. Formation and Preservation of cortical layers in slice cultures. J. Neurobiol. 23, 783-802 (1992).
- Bolz, J., Novak, N., Götz, M., Bonhoeffer, T. Formation of target-specific neuronal projections in organotypic slice cultures from rat visual cortex. Nature. 346, 359-362 (1990).
- Bolz, J., Novak, N., Staiger, V. Formation of specific afferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus. J. Neurosci. 12, 3054-3070 (1992).
- Novak, N., Bolz, J. Formation of specific efferent connections in organotypic slice cultures from rat visual cortex cocultured with lateral geniculate nucleus and superior colliculus. Eur. J. Neurosci. 5, 15-24 (1993).
- Plenz, D., Aertsen, A. Neural dynamics in cortex-striatum co-cultures. I. Anatomy and electrophysiology of neuronal cell types. Neurosci. 70, 861-891 (1996).
- Plenz, D., Aertsen, A. Neuronal dynamics in cortex-striatum co-cultures. II. Spatio-temporal characteristics of neuronal activity. Neurosci. 70, 893-924 (1996).
- Plenz, D., Kitai, S. T. Organotypic cortex-striatum-mesencephalon cultures: the nigro-striatal pathway. Neurosci. Lett. 209, 177-180 (1996).
- Plenz, D., Kitai, S. T. Up' and 'down' states in striatal medium spiny neurons simultaneously recorded with spontaneous activity in fast-spiking interneurons studied in cortex-striatum-substantia nigra organotypic cultures. J. Neurosci. 18, 266-283 (1998).
- Plenz, D., Herrera-Marschitz, M., Kitai, S. T. Morphological organization of the globus pallidus-subthalamic nucleus system studied in organotypic cultures. J. Comp. Neurol. 397, 437-457 (1998).
- Plenz, D., Kitai, S. T. A basal ganglia pacemaker formed by the subthalamic nucleus and external globus pallidus [see comments]. Nature. 400, 677-682 (1999).
- Plenz, D., Kitai, S. T. Generation of high-frequency oscillations in local circuits of rat somatosensory cortex cultures. J Neurophysiol. 76, 4180-4184 (1996).
- Klostermann, O., Wahle, P. Patterns of spontaneous activity and morphology of interneuron types in organotypic cortex and thalamus-cortex cultures. Neurosci. 92, 1243-1259 (1999).
- Cowan, R. L., Wilson, C. J. Spontaneous firing patterns and axonal projections of single cortico-striatal neurons in the rat medial agranular cortex. J. Neurophysiol. 71, 17-32 (1994).
- Porter, L. L., Rizzo, E., Hornung, J. P. Dopamine affects parvalbumin expression during cortical development in vitro. J Neurosci. 19, 8990-9003 (1999).

{kind=link}