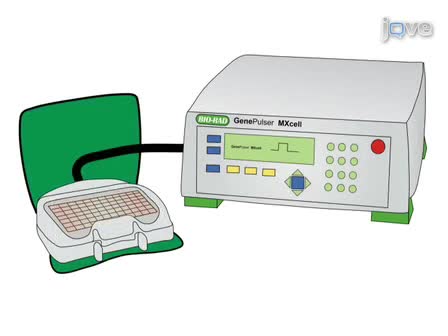
Here, we present a protocol for synthesizing virus-like particles using either baculovirus or mammalian expression systems, and ultracentrifugation purification. This highly customizable approach is used to identify viral antigens as vaccine targets in a safe and flexible manner.