When SiO2 beads are mixed with a solution of vesicles composed of phospholipids, fluorescent lipids and other lipids such as gangliosides, the vesicles rupture on the SiO2 bead surfaces to form SSLBs, as shown schematically in Figure 1a. After washing the SSLBs, a drop of SSLB solution is placed on a microwell array, and the beads are allowed to settle to the surface. (Figure 1b1) This can also be done with a suspension of phospholipid vesicles or natural membrane particles, which are shown in Figure 1b2. A fluorescence image of phospholipid vesicles adsorbed to a microwell array substrate is shown in Figure 1b3. Then the PDMS squeegee is made to slide gently across the surface to remove any SSLBs (Figure 1c1), vesicles or natural membrane particles (Figure 1c2) that did not settle into microwells. This results in a SSLB array in which each microwell contains at most one SSLB (Figure 1d1) or an array where multiple vesicles or natural membrane particles are in each microwell (Figure 1d2). An image of a microarray composed of fluorescently labeled vesicles is shown in Figure 1d3.

Figure 1. Formation of SSLBs and SSLB array assembly. (a) Illustration of a SSLB composed of 3 types of lipids on an SiO2 bead: egg PC (black), Rho-DPPE (red) and GM1 (blue). (b1-d1) Process flow for assembly of a SSLB array showing SSLBs scattered on a microwell array (b1), use of a PDMS squeegee to clear the surface of SSLBs not settled into microwells (c1), and the final SSLB array (d1). (b2-d2) The same process illustrated for the case of phospholipid vesicles or natural membrane particles where more than one vesicle or particle can be accommodated in each microwell. (b3) A fluorescence image of phospholipid vesicles adsorbed onto a microwell substrate corresponding to the illustration in (b2). (d3) A fluorescence image of a microarray composed of phospholipid vesicles corresponding to the illustration in (d2). Adapted from references 20 and 21 and used with permission of the American Chemical Society.
Individual SiO2 beads supports for SSLBs in microwells were imaged with scanning electron microscopy (SEM) from both the angled top-view and cross-sectional perspectives. A top view of an area approximately 15 µm x 15 µm is shown in Figure 2a, while Figure 2b shows a cross-sectional view of a single bead immobilized in a microwell. The lighter layer coating the microwell is 100 nm of Al2O3 deposited by atomic layer deposition to tune the well diameter so that the wells are capable of accommodating only single beads.

Figure 2. Scanning electron microscopy images of SSLB arrays. (a) An electron micrograph of a 15 mm x 15 mm area showing SSLB bead supports immobilized in microwell arrays. (b) A single 700 nm diameter SSLB in a microwell. The microwell coating is Al2O3, which is deposited to shrink the microwell so that only individual beads can fit inside. Adapted from reference 20 and used with permission of the American Chemical Society.
Once the SSLB arrays are prepared, they can be imaged by fluorescence microscopy. Figure 3a shows a 100 µm × 100 µm area of an array with 936 SSLBs. To determine the percent of occupancy of an array, the fluorescence image can be compared with a brightfield image of the same area. Figure 3b shows an area with 550 microwells and 416 SSLBs for an occupancy rate of 76%.
Sequential deposition of different types of SSLBs was used to create arrays with more than one type of SSLB, where the identity of individual SSLBs was determined by the absence or presence of a toxin receptor (GM1) and its binding of cholera toxin (CTx). The assembly process was the same as for a single SSLB type but was carried out a second time with a different SSLB identity. The starting concentration of SSLBs was 10x lower to reduce occupancy of the first type of SSLB and to allow more vacancies to be occupied by the second type of SSLB. In the Figures 3c-3e, an SSLB array with two types of SSLBs is shown. All of the SSLBs contain red fluorescent Rho-DPPE (Figure 3c), but only some of the SSLBs contain GM1, a ganglioside that is the receptor for the B-subunit of CTx. When exposed to green fluorescent Alexa 488-conjugated CTx, only the SSLBs containing GM1 fluoresce in the green channel (Figure 3d). When the red and green images are merged, it is possible to determine which SSLBs contain GM1 (yellow) and which do not (red) (Figure 3e).

Figure 3. Fluorescence imaging of SSLB arrays. (a) A 100 µm x 100 µm area containing 936 Rho-DPPE-labeled egg PC SSLBs. (b) A brightfield and fluorescence overlay showing a SSLB with 76% of the microwells occupied. (c-e) A SSLB array containing SSLBs that contain Rho-DPPE (c), a fraction of which also contain GM1, which is bound by Alexa-488-conjugated CTx (d). (e) Merger of red and green channels where colocalized fluorescence indicates the presence of GM1 on the SSLB. The blue circles indicate SSLBs that only fluoresce red, i.e. lacking GM1. Adapted from reference 20 and used with permission of the American Chemical Society.
Fluorescence data from SSLB arrays was analyzed by measuring the intensity of individual SSLBs in an image. The SSLB intensity data from arrays were compiled into histograms. Figure 4a shows one such histogram from the analysis of Rho-DPPE fluorescence from a SSLB array composed solely of SSLBs containing egg PC and 1 mol% Rho-DPPE. The data in Figure 4a were acquired by analyzing the image in Figure 3a. We found the SSLB arrays to be robust and stable for at least one week when refrigerated and submerged in buffer. The arrays did not lose any fluorescence intensity, nor did the occupancy of the arrays change over the course of one week (Figure 4b).

Figure 4. (a) Histogram showing the distribution of fluorescence intensities for the SSLB array shown in Figure 3a. The mean intensity for each SSLB in the array was used to compile the histogram. The solid line is a Gaussian fit to the data. (b) Mean array occupancy (black circles) and fluorescence intensity (red squares) for a SSLB array monitored over the course of one week. Adapted from reference 20 and used with permission of the American Chemical Society.
We made arrays composed of SSLBs with varying concentrations of GM1 (0, 0.5, 1 and 2 mol%) and exposed them to a fixed concentration of CTx as well as arrays with SSLBs with concentrations of GM1 and exposed them to varying concentrations of CTx. The later experiment was used to determine the equilibrium dissociation constant (Kd) for GM1/CTx binding. The SSLB arrays with varying GM1 concentration were exposed to 50 nM Alexa-488 labeled CTx for 1 hr then washed and imaged. The intensity histograms for SSLB arrays with 0.5 – 2 mol% GM1 exposed to 50 nM Alexa 488-conjugated CTx can be seen in Figure 5a. Figure 5b shows the linear dependence of mean fluorescence intensity upon GM1 concentration. When the concentration of GM1 in SSLBs is fixed at 2 mol% and the SSLB arrays are exposed to CTx ranging from 16 pM to 158 nM, the binding response follows sigmoidal curves as shown in Figure 5c. The curves fit to this data are based upon two different binding models: the Langmuir isotherm (Eq. 1) and the Hill-Waud model (Eq. 2). Please click here to view a larger version of this figure.
 (1)
(1)
 (2)
(2)
Where F is the fluorescence intensity at a given CTx concentration, Fmax is the fluorescence intensity when binding is saturated, [CTx] is the CTx concentration, KD and KH are the dissociation constants from the Langmuir and Hill-Waud fits, respectively, and n is cooperativity coefficient in the Hill-Waud model. The cooperativity coefficient (n) estimates the amount of multivalent binding, where an n greater than one indicates that each CTx molecule binds more than one GM1. The concentration of CTx at 0.5 x Fmax is the KD or KH for the GM1/CTx interaction. In our experiments we calculated a KD of 1.6 ± 0.2 nM and KH of 1.4 ± 0.2 nM with an n value of 1.3, indicating cooperative binding. The dissociation constants for the GM1/CTx interaction vary widely in the literature, ranging over 5 orders of magnitude from 4.5 pM to 370 nM26,27.
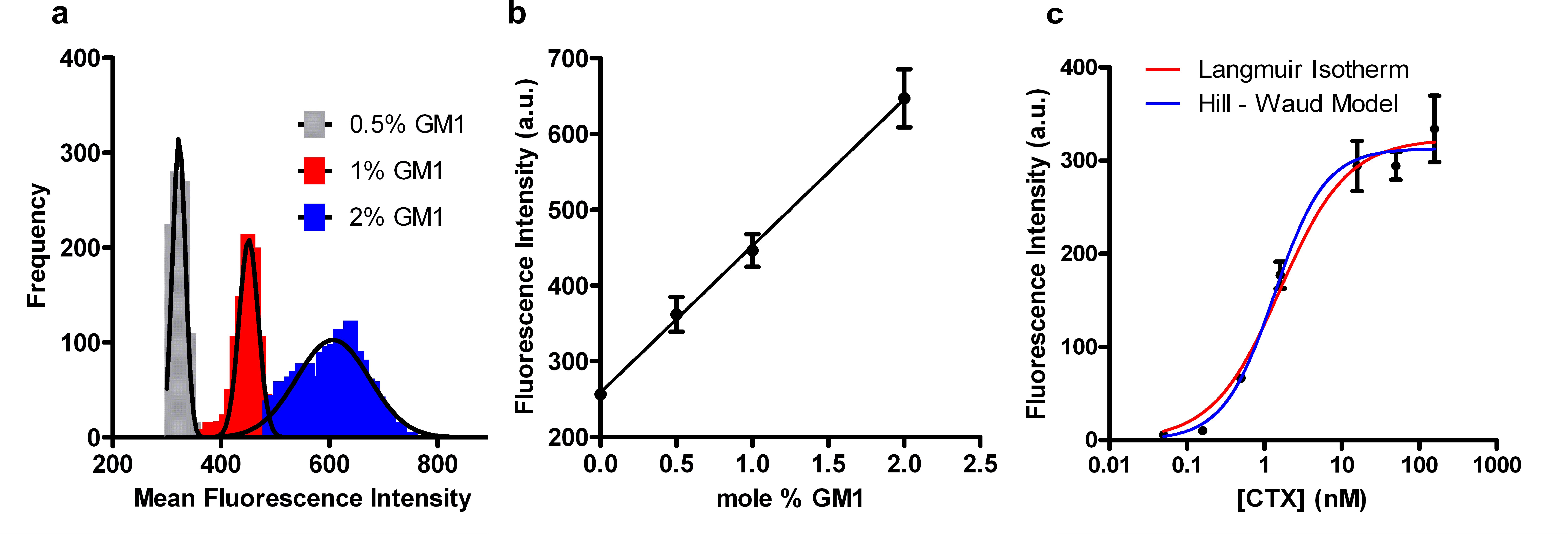
Figure 5. CTx/GM1 binding assays on SSLB arrays. (a) Histograms of the fluorescence intensity of individual SSLBs within SSLB arrays. The arrays contained SSLBs with 0.5, 1 or 2 mol% GM1 and were exposed to 50 nM Alexa 488-conjugated CTx. The solid lines are Gaussian fits. (b) Plot of the distribution means from (a) as a function of GM1 concentration. (c) Binding curves showing the distribution means from arrays containing SSLBs with 2 mol% GM1 after exposure to Alexa-488-conjugated CTx varying from 16 pM to 158 nM. The lines are fits to the Langmuir isotherm (red) and Hill-Waud (blue) binding models. The error bars in (b-c) represent the standard deviations of the distributions of fluorescence intensity corresponding to each data point. Adapted from reference 20 and used with permission of the American Chemical Society.
As mentioned earlier, it is also possible to form arrays with biomembrane material derived from natural sources21. The arrays are formed the same way, by dispersing membrane particles on a microwell substrate, then cleaning the top surface with the squeegee to leave behind membrane particles in the microwells. Figure 6 shows examples of myelin and neuronal lipid raft microarrays. In Figure 6a, myelin particles are labeled with the lipophilic fluorophore FM1-43. Lipid rafts are known to be enriched in cholesterol and gangliosides such as GM1; thus, Alexa 488-conjugated CTx binds strongly to lipid raft microarrays as shown in Figure 6b, while fluorescently labeled streptavidin (SAPE) does not bind to the array. (Figure 6c) We also created stripe arrays by delivering myelin and lipid raft particles to the microwell substrate with a microfluidic chip with 250 µm-wide channels. After particle delivery, the microfluidic chip was detached from the microwell substrate and the squeegee process carried out as usual. The stripe arrays were then exposed to a fluorescent anti-oligodendrocyte IgM antibody (IgM O4), which binds sulfatide found in myelin. (Figure 7) The myelin and lipid raft stripe microarrays in Figure 7 are shown at increasing levels of magnification, and at the highest level of magnification, it is readily apparent that IgM O4 only binds to the myelin microarrays because sulfatide is absent from the lipid rafts.

Figure 6. Myelin particle and neuronal lipid raft particle arrays. (a) A myelin particle microarray where the myelin particles are rendered fluorescent by the lipophilic fluorophore FM1-43. The dashed line indicates the edge of the array area. (b) Lipid raft microarray showing CTx binding to the lipid raft particles. (c) Lipid raft microarray exposed to fluorescent streptavidin-phycoerythrin (SAPE) showing little to no nonspecific binding. Adapted from reference 21 and used with permission of the American Chemical Society.

Figure 7. Multicomponent arrays formed by microfluidic delivery of myelin and neuronal raft membranes. (a) Fluorescence image after myelin and rafts were deposited via microfluidic channels on the microarray substrate. The left and right stripes contain myelin and the middle stripe contains neuronal raft membranes. The membranes are stained with FM1-43, which labels all lipid material. The scale bar is 250 μm. (b) Fluorescence image of the same three stripes after applying the PDMS squeegee and incubating with IgM O4. The scale bar is 250 μm. (c-e) Magnified images from the three stripes showing that IgM O4 only binds to the myelin microarrays: (c) left stripe and (e) right stripe. The scale bars in panels c-e are 30 μm. Adapted from reference 21 and used with permission of the American Chemical Society.
