

Summary
Описан весьма параллельный метод измерения специфического участка расщепления ДНК на уровне одной молекулы. Этот протокол демонстрирует технику с использованием ограничения endonuclease NdeI. Метод может быть легко изменен для изучения любого процесса, что приводит к сайту конкретных расщепления ДНК.
Abstract
Расщепление ДНК (SSDC) является ключевым шагом во многих клеточных процессах, и это имеет решающее значение для редактирования генов. Эта работа описывает кинетический анализ, способный измерять SSDC во многих отдельных молекулах ДНК одновременно. Привязные бисером субстратные ДНК, каждый из которых содержит одну копию целевой последовательности, готовятся в канале микрофлюидного потока. Внешний магнит применяет слабую силу к парамагнитным бусинкам. Целостность до 1000 отдельных ДНК можно контролировать с помощью визуализации микробусов под темным полем изображения с помощью широкого поля, с низким увеличением цели. Инъекция эндонуклеазы ограничения, NdeI, инициирует реакцию расщепления. Видео микроскопия используется для записи точного момента каждого расщепления ДНК, наблюдая кадр, в котором связанный шарик движется вверх и из фокусной плоскости цели. Подсчет бисера кадр за кадром количественно определяет реакцию, и экспоненциальная подгонка определяет скорость реакции. Этот метод позволяет сбор количественных и статистически значимых данных о реакции одной молекулы SSDC в одном эксперименте.
Introduction
Расщепление ДНК(SSDC) является ключевым шагом во многих геномных транзакциях. Например, системы бактериального ограничения модификации (RM)1 и CRISPR2 защищают клетки от атак фагов и плазмидов, распознавая и расщепляя иную ДНК в определенных последовательностях. В типе II RM, ограничения эндонуклеи (REs) распознают короткие 4-8 базовых пар (bp) последовательности через белково-нуклеиновой кислотывзаимодействий 3. СВЯЗАННЫе с CRISPR эндонуклеи, такие как Cas9, связываются с сайтами через гибридизацию целевого сайта с crRNAs, связанными с эндонуклеями4. Создание сайта конкретных двойных мель перерывов (DSBs) также являются первым шагом во многих событиях рекомбинацииДНК 5. Например, разнообразие антигенных связывающих областей, созданных рекомбинацией V(D)J, требует распознавания и расщепления конкретных целевыхобъектов 6. Некоторые транспозоны, как известно, нацелены на конкретные последовательности ДНК, атакже 7. Неудивительно, что многие ядра, связанные с конкретными участками, участвующие в этих процессах, такие как Cas9, являются ключевым компонентом технологий редактирования генов8. Кроме того, для редактирования геномов были также разработаны новые эндонуклеазы для конкретныхсайтов (т.е. ядра цинковых пальцев 9 и TALENS10).
Многие методы были использованы для измерения кинетики сайта конкретных расщепления нуклеиновых кислот. К ним относятся гель анализ, флуоресценция11,12, и секвенирования на основеметодов 13. Значительный прогресс был достигнут с привязыванием микробусов, что позволяет DSBs в отдельных молекул ДНК, которые будут обнаружены движением шарика после разделения нити. В этих методах, различные типы сил используются для обеспечения разделения нити и движения шарика после расщепления. В одном случае, оптические ловушки были использованы для измерения расщепления ДНК EcoRV14. В этих экспериментах целевой поиск является целью исследования, при этом условия оптимизированы таким образом, чтобы привязкой к конкретному сайту был шаг ограничения скорости. Одним из недостатков оптических ловушек является то, что только одна ДНК может наблюдаться одновременно. Кроме того, периодические большие силы потянув должны быть применены для проверки на разделение нити.
Другой метод использует сочетание потока и слабых магнитных сил, чтобы тянуть на шарик в непрерывной манере15. Таким образом, диффузия ограниченного расщепления NdeI измеряется. Используемый метод позволяет одновременно измерять сразу несколько сотен ДНК, что позволяет достичь статистической значимости в ходе одного эксперимента. Эксперименты на основе магнитных пинцетов также были использованы. В одном из таких исследований, ретровирусной интегразы был изучен в том числе DSB в вставке олигонуклеотид16. Успешная интеграция привела к включению DSB в привязанную ДНК и потере прикрепленного шарика. В аналогичном исследовании АТФ-зависимого типа III ограничения эндонуклеазы EcoPI, десятки ДНК наблюдались в одном эксперименте17. Магнитные пинцеты имеют то преимущество, что напряжение, а также цикл ДНК, можно контролировать и контролировать во время реакции.
Здесь представлен весьма параллельный метод одной молекулы для измерения кинетики SSDC, который использует недавние улучшения в крупномасштабных привязываниях ДНК. Этот метод является улучшение и расширение предыдущих методов, используемых дляизмерения репликации ДНК 18, длинаконтура ДНК 19, и расщепление REs15. В этом методе линейные ДНК, содержащие одну копию последовательности распознавания, готовятся с биотином на одном конце и дигоксигенином на другом. Биотин связывает стрептавидин, который ковалентно крепится к парамагнитной микробусу. ДНК-бисерные комплексы вводятся в микрофлюидный канал, который был функционализирован с анти-digoxigenin FAB фрагментов. Затем ДНК привязывается к точкам крепления поверхности через привязку дигоксигенина к адсорбированным фрагментам FAB. Слабые магнитные силы, применяемые с помощью постоянного магнита, не держат шарик от прилипания некондиентного к поверхности. Образцы могут быть введены быстро (Lt;30 s) в канал потока, чтобы активировать реакцию расщепления. Поток отключается во время сбора данных. Поскольку каждая ДНК расщепляется, точное время расщепления может быть определено путем записи кадра, в котором шарик движется вверх и из координационного плана цели, тем самым исчезая из видеозаписи. Для количественной оценки прогресса реакции можно использовать рамку за кадром, подсчитывая оставшиеся бусы.
Ниже приведен полный протокол, а также пример данных, собранных с помощью NdeI. В качестве примера того, как метод может быть применен, расщепление ставки для целого ряда концентраций белка измеряются в двух различных концентрациях магния, существенного кофактора металла. Хотя это применение протокола использует NdeI, метод может быть адаптирован для использования с любым сайтом конкретных нуклеазы путем изменения конструкции субстрата ДНК.
Subscription Required. Please recommend JoVE to your librarian.
Protocol
1. Создание ячейки потока
- Мытье крышки
- Место coverslips в окрашивание банки и sonicate с этанолом (EtOH), а затем с 1 M KOH (по 30 мин каждый). Чтобы избежать осадков KOH в EtOH, тщательно промойте ddH2O между мойки.
- Повторите как EtOH и KOH стиральные шаги 1x для общего числа четырех моет (два EtOH и два KOH). Храните очищенные крышки в ddH2O в окрашивающих банках.
- Вырезать погрузки и выхода труб (8 см в длину) с помощью чистой бритвой и вставить в отверстия в чистом стеклянном слайде. Используйте PE-20 для входов и PE-60 для розетки. Epoxy в течение 5 минут, чтобы обеспечить трубки и обрезать любые избыточные трубки.
ПРИМЕЧАНИЕ: Стеклянные слайды измеряют 2'' x 3'' x 1 мм. Отверстия просверлены парами на 15 мм друг от друга. Каждая пара формирует любой конец одного канала. Трубка с меньшим идентификатором используется для входе, так как это уменьшает мертвый объем вверх по течению и, таким образом, время смешивания требуется (Рисунок 1). - Выстроиться и применить precut двустороннюю ленту с каналом шаблон вырезать над отверстиями на стеклянной горке. Гладкая с пластиковыми типсами для достижения хорошего уплотнения.
ПРИМЕЧАНИЕ: Двусторонняя лента, используемая в этом эксперименте, имеет толщину 120 мкм, а каналы шириной 2 мм и длиной 15 мм. Каналы вырезаются с помощью ножапринтера (Таблица материалов). Можно поместить до четырех каналов на одном обложке. Смотрите рисунок 1 для изображения ячейки потока. - После отслаивания от поддержки, нанесите чистый крышку (высушенную сжатым воздухом) на ленту и снова разглайте пластиковыми типсами для хорошего уплотнения.
- Epoxy края от крышки, чтобы запечатать поток ячейки и дайте ему вылечить.
2. Подготовка помеченной ДНК для привязывания
- В ПЦР-трубке подготовьте 50 МКЛ смеси реакции ПЦР, содержащей полимеразу ДНК высокой точности 0,02 U/L, 200 МКМ dNTPs, 0,5 МКМ вперед грунтовки, 0,5 МКМ обратной грунтовки, и 250 нг M13mp18 векторной ДНК.
ПРИМЕЧАНИЕ: Здесь, вперед грунтовки (биотин-CCAACTTAATCGCCTTGC) и обратной грунтовки (digoxigenin-TGACCATTAGATACATTTCGC) были выбраны для усиления региона около 1000 bp длиной, охватывающих от позиций 6338 до 107 в круговом геноме M13mp18 ДНК. В центре усиленной области имеется единый сайт NdeI. Перемотка вперед 5ʹ помечена дигоксигенином, который связывает анти-дигоксигенин на крышке. Обратная грунтовка 5ʹ помечена биотином, который связывает бисер со стрептавидиным покрытием. - Вставьте трубку ПЦР в термоциклер и следуйте циклу, как показано в таблице 1.
- Очистите продукт PCR с помощью комплекта очистки ПЦР в соответствии с протоколом производителя.
ПРИМЕЧАНИЕ: Использование комплекта, указанного в таблице материалов, типичный выход ДНК составляет 2 мкг.
3. Привязка ДНК и бисера
- Приготовьте 10 мл буфера A (1 M Tris-HCl (pH ) 7,5 , 50 мМ НаКл, 2 мМ МгКл2, 1 мг/мл β-Casein, 1 мг/мл Плуронический F-127). Дега в вакуумном децикаторе не менее 1 ч.
- Для функционализации ячейки потока в канал потока вводится 25 МКЛ антидигоксигенина ФРАГМЕНТОВ ФАБ (20 мкг/мл) в PBS. Используйте гель загрузки советы, чтобы вписаться в PE-60 труб. Инкубация при комнатной температуре (RT) в течение 30 мин.
- После инкубации смойте канал, потянув 0,5 мл буфера А через канал с помощью шприца. Позаботьтесь о том, чтобы не вводить воздух в канал.
- После функционализации намонтировать ячейку потока на перевернутом микроскопе. Подключи розетку к шприц-насосу и поместите входную трубку в микроцентрифугную трубку, содержащую буфер А.
- Вручную потяните не менее 0,5 мл буфера А, чтобы промыть систему и загрунтовать насос. Пусть насос работает на 10 йл/ мин, по крайней мере 5 минут, чтобы уравночные системы.
- Для подготовки бисера(Таблица материалов), вихрь запас бутылку бисера и пипетки 1,6 л из 10 мг / мл бульона бусы в 50 л буфера, то вихрь.
- Используя магнитный сепаратор, пипетка из буфера и resuspend в 50 йл буфера, то вихрь.
- Повторите шаг 3.7 2x в общей сложности три моет. Для последней стирки, перерасход в 100 мкл буфера и вихря для достижения окончательной концентрации 160 мкг/мл.
- Чтобы упраз сложности ДНК и бисера, сначала подготовьте 480 МКЛ 0,5 рМ с маркировкой ДНК-субстрата в буфере А. Затем, пипетка в 20 йл 160 мкг / мл шарик подвески, убедившись, что вихрь бисера перед пипеткой. Поместите на ротатор в течение 3 мин.
- Через 3 мин. немедленно загружайте в канал со скоростью потока 10 мл/мин в течение 15 мин или до тех пор, пока не будет соблюдено достаточное количество привязывания бисера.
ПРИМЕЧАНИЕ: Бисер не должен быть настолько плотно упакован, что они взаимодействуют друг с другом на поверхности (см. раздел обсуждения). - Чтобы вымыть канал всех свободных бусин, переключите входную трубку на свежую трубку буфера А и втмитесь со стороны 50 йл/мин в течение по крайней мере 10 минут или до тех пор, пока не будут замечены свободные бусы.
4. Сбор и анализ данных
- Чтобы подготовиться к сбору данных, поместите входную трубку в микроцентрифугную трубку, содержащую не менее 100 МКЛ NdeI (0,25-4,00 U/mL) в буфере A. Нижняя постоянная магнит над каналом потока, и располагаем источник света от оси для изображения темного поля.
ПРИМЕЧАНИЕ: Два кольцеобразных редкоземельных магнита, эпоксидные вместе, находятся на 8 мм над активной поверхностью канала потока с помощью кантилеверированного оптического столба во время сбора данных. Гусиная лампа шеи используется для внеосевого светового курса. - Используйте коммерческий микроскоп, видеокамеру и программное обеспечение для сбора данных(Таблица материалов). В программном обеспечении щелкнитевкладку«Экспозиция» и установите«Время экспозиции»до 10 мс. Нажмитена вкладку Timelapseи установите "Image Count" до 600, "Длительность" до 20 мин, и "Интервал" до 2 с. Нажмитекнопку"Беги", чтобы начать сбор данных.
- На шприц-насосе установите скорость потока до 150 мкл/мин и объем впрыска до 80 йл. Нажмите "Run" на 1 мин в сборе данных. После инъекции выключите насос и закройте клапан, чтобы предотвратить поток во время сбора данных.
- После сбора данных открытое программное обеспечение для анализа изображений(Таблица материалов). Подвкладкой "Файл",выберите" Импорт " "Последовательность изображений". Найдите файлы изображений во всплывающем меню и нажмите кнопку "Открыть".
- Установите порог, выбрав "Adjust Theshold" под "Изображение "выдвижной меню. Используйте бар ползунка, чтобы установить пороговое значение для определения ярких пятен, соответствующих бисеру на изображении.
- Подсчитайте яркие пятна в каждом кадре, нажавкнопку «Анализчастиц» в«Анализе»выдвижного меню. Нажмите "OK", а затем выберите" Да", чтобы обработать все изображения. Сохраните файл результатов.
ПРИМЕЧАНИЕ: Это позволит сохранить файл данных, который содержит количество бусинок в каждом записанном видеорамку. - Программное обеспечение для анализаоткрытых данных (Таблицаматериалов) и импортировать файл результатов, нажав на " Импорт изтекстовогофайла " в "Файл" выдвижной меню. Участок данных подсчета шарика против времени.
- Fit количество бисера против времени, нажав "Кривая Fit" в "Анализ "выдвижной меню. Выберитеуравнение «Естественный экспонент»и нажмите кнопку «Try Fit » "Хорошо".
ПРИМЕЧАНИЕ: В подходящий регион должны быть включены только данные, записанные после введения выборки. Подходящим параметром в экспоненте фитинговой функции будет скорость расщепления.
Subscription Required. Please recommend JoVE to your librarian.
Representative Results
Используя этот метод, SSDC ставки NdeI были измерены для диапазона концентраций белка (0,25-4,00 U/mL) в двух различных концентрациях магния (2 мМ и 4 мм). Каждое из этих условий было воспроизведено по крайней мере дважды, с несколькими сотнями до 1000 привязанных ДНК за эксперимент. Рисунок 2 описывает экспериментальный дизайн. На рисунке 3 приведены примеры данных сбора и анализа. Рисунок 4 иллюстрирует, как скорость зависит от концентрации белка в двух концентрациях магния. Можно засзнать, что при достаточно низких концентрациях белка скорость пропорциональна белку и не зависит от магния. Для достаточно высоких концентраций белка, скорость зависит от магния, но независимо от концентрации белка.
| Шаг | Описание | Температура (КК) | Время (ы) |
| 1 | Денатурация | 98 | 30 |
| 2 | Расплава | 98 | 10 |
| 3 | Аннеал | 60 | 30 |
| 4 | Расширить | 72 | 30 |
| 5 | Окончательное продление | 72 | 120 |
Таблица 1: Параметры ПЦР. Показаны температуры и продолжительность шагов программы термоциклера, используемых в шаге 2.2 протокола. Расплав, аннеал и удлинить шаги (шаги 2, 3 и 4) повторяются 30x.
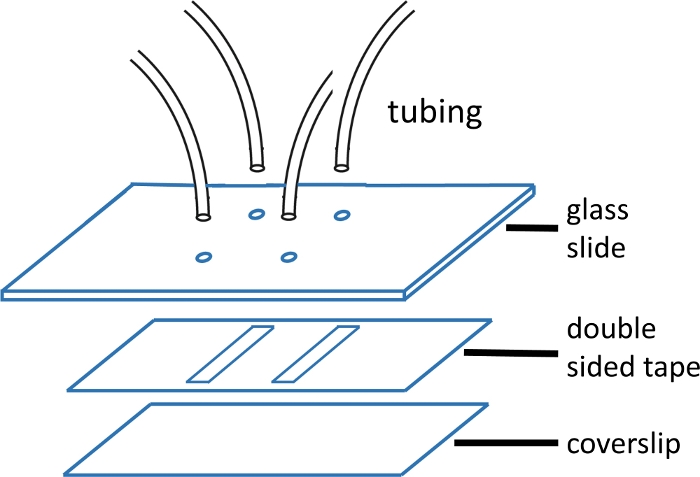
Рисунок 1: Микрофлюидный поток клеточной конструкции.
Верхний стеклянный слайд (2'' x 3', толщиной 1 мм) предварительно просверлен отверстиями, соответствующими шаблону канала. Входные и розетки трубки вставляются в отверстия и фиксируется с эпоксидной смолой до крепления ленты и крышки стекла. Двусторонняя лента предварительно вырезана с узором канала. Нижняя горка (#1 или #1,5 крышки) предварительно очищается с помощью протокола, описанного в основном тексте. После сборки край крышки стекла запечатывается эпоксидной смолой. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.

Рисунок 2: Экспериментальный дизайн.
(A)Метод привязывания ДНК. Двухместная ДНК (1 кбп), помеченная дигоксигенином на 5ʹ, прикрепляется к поверхности клетки потока через антидигоксигенин-дигоксигенин взаимодействия. ДНК 3ʹ, помеченный биотином, крепится к микробусу через стрептавидин-биотин. Участок расщепления NdeI расположен в центре ДНК. (B)Экспериментальная установка во время сбора данных, показывающая магнит и объективное положение. Постоянный магнит поддерживает слабую восходящую силу на шарике во время реакции расщепления. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
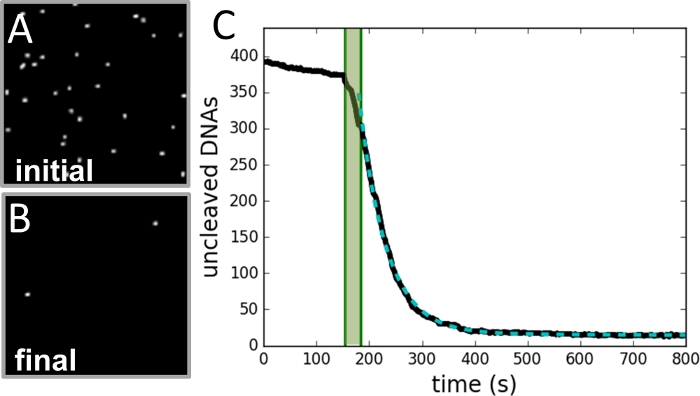
Рисунок 3: Пример сбора и анализа данных.
(A) Изображение области бисера, принятых до расщепления реакция инициируется. (B)Изображение того же региона после завершения реакции. (C) Участок количества бисера против времени (черная кривая), как определено из каждого кадра видеозаписи. Затененная зеленая зона отмечает период инъекций фермента и не входит в пригонку. Данные подходят к одной экспоненциальной кривой (dashed green curve), с постоянной скоростью распада, равной скорости реакции. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.

Рисунок 4: Расщепление NdeI зависит от концентрации белка и магния.
Участок измеренной скорости SSDC NdeI для ряда концентраций белка в двух различных концентрациях магния: 2 мМ (синие круги) и 4 мМ (зеленые квадраты). Бары ошибок представляют SEM. Разбитые кривые являются линиями тренда и не подходят к теории. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
Subscription Required. Please recommend JoVE to your librarian.
Discussion
Протокол может быть использован для измерения кинетики любой системы SSDC, при условии, что разделение нити наблюдается в ходе эксперимента. Обнаружение расщепления зависит от наблюдения за отслоение привязали шарика и, следовательно, знаменует собой момент разделения нити. Все предыдущие шаги происходят до обнаружения расщепления; таким образом, регистрируется только общее время транзита.
Крышки клеток потока функционализируются с помощью неспецифической асорпции белка антител к чистому стеклу. Недостаточно очищенное стекло может повлиять на связывание антител. При привязывания плотность бисера должна быть достаточно низкой, чтобы бисер не взаимодействовал. Плотность поверхности точек крепления может контролироваться концентрацией антител во время функционализации. Общее количество бусин зависит от размера поля зрения. В этом случае, от нескольких сотен до 1000 привязанных бусинок было достаточно для хорошей статистики и избежать бисера бисер взаимодействия. Во время инъекции бисера, поверхностная привязка отслеживалась с помощью видео в прямом эфире. Инъекция бисера была остановлена, когда, по оценкам, количество бисера было от 500 до 1000 бусин.
Самые быстрые скорости расщепления, которые могут быть точно измерены, ограничены временем смешивания ячейки потока. Время смешивания в клетках ламинарного потока зависит от нескольких факторов. Диффузия на поверхность является ключевым шагом; таким образом, время смешивания зависит от коэффициента диффузии реакционные. Значительный сноень, который происходит в входной трубе, которая транспортируется образец в канал потока из образца резервуара, может увеличить время, необходимое для обеспечения адекватного смешивания на поверхности реакции в канале. Было установлено, что время смешивания может быть сокращено за счет сокращения мертвого объема восходящего потока и увеличения скорости потока. При внутреннем диаметре 380 мкм и максимальной длине 8 см для входных труб (и скорости потока 150 йл/мин), было установлено, что время инъекции может быть сокращено до 20 евро, не влияя на измеренную скорость расщепления. Так как время смешивания зависит от коэффициента диффузии реакционные, она должна быть определена отдельно для каждого фермента или декольте активатора изучены.
Метод привязывания допускает неспецифический разрыв троса, предположительно из-за диссоциации комплекса дигоксигенина-антитела или высвобождения антитела с поверхности. Это приводит к воспроизводимой фоновой скорости потери шарика присутствует перед инъекцией фермента в размере 3 х 10-4 с-1. Этот систематический эффект может быть исправлен либо путем вычитания фоновой ставки из измеренной скорости расщепления, либо путем моделирования фона в подходящем уравнении. Тем не менее, расщепление ставки ниже, чем эта нижняя граница будет менее надежно измеряется.
Несовершенная пассивация поверхности может привести к неправильному привязыванию. Это приводит либо к увеличению фракций «застрявших» бусинок, которые не уходят во the время эксперимента, либо к неправильно привязанным бусинкам, которые очень медленно отмежеваться от поверхности. Это создает более высокий и, возможно, наклонный базовый уровень в обработанных данных. Было установлено, что при надлежащей очистке крышки скользит и свежеприготовленные β-casein фондового решения, эти эффекты были минимальными для большинства наборов данных. Для случайных наборов данных, которые показывают это, изменение функции установки (включить наклонный базовый уровень) может исправить для этого эффекта.
Текущий протокол может быть расширен несколькими способами. Дальнейшая изоляция механистических шагов после связывания целевого участка может быть выполнена с использованием предварительно связывающего формата, в котором белок вводится при отсутствии необходимых кофакторов. Эта идея тестируется путем введения NdeI в отсутствие магния. В этих условиях белок связывается со своим местом копята, но не расщепляет ДНК. Инъекция магния после этого связывающего шага активирует расщепление, что приводит к быстрой потере бисера. activates Экспериментальная установка также позволяет контролировать конформацию и напряжение ДНК, изменяя конфигурацию магнита или добавляя поток. При низких силах в этих экспериментах, ДНК становится частично спиральный. Изменение сил немного может иметь драматическое влияние на конформацию ДНК. Например, в буферных условиях, в которых целевой поиск является ограничение скорости, изменение конформации ДНК может проверить на влияние прыжков на целевой поиск. В буферных условиях, в которых шаг гидролиза является ограничение скорости, изменение силы может зондировать влияние напряжения ДНК на гидролиз фосфодиестер облигаций. Следует отметить, что при низком увеличении эти конформациальные изменения не могут наблюдаться. В результате небольшие движения в положении шарика должны быть отслежены под более высоким увеличением, чтобы проверить, что конформация ДНК может контролироваться.
Анализ данных может быть расширен различными способами. Эта работа применяет простой метод подсчета бисера с последующим кривой установки одной экспоненциальной функции. Методы, основанные на анализе времени проживания могут быть использованы, атакже 20. Распределение времени проживания отдельных ДНК может быть проанализировано с помощью кривой установки или с помощью более сложных методов, таких как обобщенный методмоментов 21.
Subscription Required. Please recommend JoVE to your librarian.
Disclosures
У авторов нет конфликта интересов, чтобы раскрыть.
Acknowledgments
Эта работа была поддержана Грантом Национального научного фонда MCB-1715317.
Materials
| Name | Company | Catalog Number | Comments |
| 5 minute Epoxy | Devcon | 14250 | |
| anti-digoxigenin FAB fragments | Roche Diagnostics | 11214667001 | |
| camera and software | Jenoptik | GRYPHAX SUBRA | |
| data analysis software | Vernier Inc. | LP | |
| double sided tape | Grace Biolabs | SA-S-1L | |
| Dulbeccos Phosphate Buffered Saline | Corning | 21-031-CV | |
| ethanol 95% | VWR | 89370-082 | |
| forward primer: digoxigenin-CCAACTTAATCGCCTTGC | Integrated DNA Technologies | n/a | |
| image analysis software | National Institutes of Health | ImageJ | |
| inverted microscope | Nikon | TE2000 | |
| knife printer | Silhouette | ||
| M13mp18 DNA | New England Biolabs | N4040S | |
| MyOne streptavidin beads | Thermo Fisher Scientific | 65601 | |
| NdeI enzyme | New England Biolabs | R0111S | |
| PCR cleanup kit | Qiagen | 28104 | |
| pluronic F-127 | Anatrace | P305 | |
| polyethylene tubing PE-20 | BD Intramedic | 427406 | |
| polyethylene tubing PE-60 | BD Intramedic | 427416 | |
| Q5 Mastermix | New England Biolabs | M0492S | |
| rare earth magnet 0.5" OD 0.25" ID | National Imports | NSN0814 | |
| rare earth magnet 0.75" OD 0.5" ID | National Imports | NSN0615 | |
| reverse primer: biotin-TGACCATTAGATACATTTCGC | Integrated DNA Technologies | n/a | |
| syringe pump | Kent Scientific | Genie Plus | |
| β-Casein from bovine Milk | Sigma-Aldrich | C6905 |
References
- Tock, M. R., Dryden, D. T. The biology of restriction and anti-restriction. Current Opinion in Microbiology. 8 (4), 466-472 (2005).
- Garneau, J. E., et al. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature. 468 (7320), 67-71 (2010).
- Pingoud, A., Fuxreiter, M., Pingoud, V., Wende, W. Type II restriction endonucleases: structure and mechanism. Cellular and Molecular Life Sciences. 62 (6), 685-707 (2005).
- Sternberg, S. H., Redding, S., Jinek, M., Greene, E. C., Doudna, J. A. DNA interrogation by the CRISPR RNA-guided endonuclease Cas9. Nature. 507 (7490), 62-67 (2014).
- Sadowski, P. D. Site-specific genetic recombination: hops, flips, and flops. The FASEB Journal. 7 (9), 760-767 (1993).
- Schatz, D. G. Antigen receptor genes and the evolution of a recombinase. Seminars in Immunology. 16 (4), 245-256 (2004).
- Craig, N. L. Tn7: a target site-specific transposon. Molecular Microbiology. 5 (11), 2569-2573 (1991).
- Gori, J. L., et al. Delivery and Specificity of CRISPR/Cas9 Genome Editing Technologies for Human Gene Therapy. Human Gene Therapy. 26 (7), 443-451 (2015).
- Urnov, F. D., Rebar, E. J., Holmes, M. C., Zhang, H. S., Gregory, P. D. Genome editing with engineered zinc finger nucleases. Nature Reviews Genetics. 11 (9), 636-646 (2010).
- Joung, J. K., Sander, J. D. TALENs: a widely applicable technology for targeted genome editing. Nature Reviews Molecular Cell Biology. 14 (1), 49-55 (2013).
- Alves, J., Urbanke, C., Fliess, A., Maass, G., Pingoud, A. Fluorescence stopped-flow kinetics of the cleavage of synthetic oligodeoxynucleotides by the EcoRI restriction endonuclease. Biochemistry. 28 (19), 7879-7888 (1989).
- Deng, J., Jin, Y., Chen, G., Wang, L. Label-free fluorescent assay for real-time monitoring site-specific DNA cleavage by EcoRI endonuclease. Analyst. 137 (7), 1713-1717 (2012).
- Becker, W. R., et al. High-throughput analysis reveals rules for target RNA binding and cleavage by AGO2. Molecular Cell. 75 (4), 741-755 (2019).
- vanden Broek, B., Lomholt, M. A., Kalisch, S. M., Metzler, R., Wuite, G. J. How DNA coiling enhances target localization by proteins. Proceedings of the National Academy of Sciences. 105 (41), 15738-15742 (2008).
- Gambino, S., et al. A single molecule assay for measuring site-specific DNA cleavage. Analytical Biochemistry. 495, 3-5 (2016).
- Jones, N. D., et al. Retroviral intasomes search for a target DNA by 1D diffusion which rarely results in integration. Nature Communications. 7, 11409 (2016).
- van Aelst, K., et al. Type III restriction enzymes cleave DNA by long-range interaction between sites in both head-to-head and tail-to-tail inverted repeat. Proceedings of the National Academy of Sciences. 107 (20), 9123-9128 (2010).
- Williams, K., et al. A single molecule DNA flow stretching microscope for undergraduates. American Journal of Physics. 79 (11), 1112-1120 (2011).
- Song, D., et al. Tethered particle motion with single DNA molecules. American Journal of Physics. 83 (5), 418-426 (2015).
- Etson, C. M., Todorov, P., Walt, D. R. Elucidating Restriction Endonucleases Reaction Mechanisms via Dwell-Time Distribution Analysis. Biophys Journal. 106 (2), 22 (2014).
- Piatt, S., Price, A. C. Analyzing dwell times with the Generalized Method of Moments. PLoS One. 14 (1), 0197726 (2019).
